Extremozymes vs. Conventional Microbial Enzymes: A Comparative Analysis of Stability for Biomedical Applications
This article provides a comprehensive comparative analysis of the structural stability and functional resilience of extremophile-derived enzymes (extremozymes) against their conventional microbial counterparts.
Extremozymes vs. Conventional Microbial Enzymes: A Comparative Analysis of Stability for Biomedical Applications
Abstract
This article provides a comprehensive comparative analysis of the structural stability and functional resilience of extremophile-derived enzymes (extremozymes) against their conventional microbial counterparts. Tailored for researchers, scientists, and drug development professionals, it explores the foundational biochemical adaptations, details advanced methodologies for discovery and production, addresses key challenges in bioprocessing, and validates performance through direct comparative studies. The synthesis underscores the immense potential of extremozymes to overcome limitations in industrial biocatalysis and pharmaceutical manufacturing, offering robust, efficient, and sustainable solutions for demanding processes.
Defining Stability: Structural and Functional Adaptations in Microbial and Extremophile Enzymes
Enzyme stability is a fundamental property determining the efficacy, shelf-life, and economic viability of biocatalysts in industrial and pharmaceutical processes. It refers to an enzyme's ability to maintain its structural integrity and catalytic function under a variety of challenging conditions, including extreme temperatures, pH fluctuations, organic solvents, and mechanical shear [1]. For researchers and drug development professionals, understanding and enhancing enzyme stability is paramount for developing efficient bioprocesses and therapeutic enzymes.
The study of enzyme stability has been profoundly advanced by investigating organisms that thrive in extreme environments, known as extremophiles. These organisms have evolved unique molecular adaptations that allow their enzymes, termed extremozymes, to function under conditions that would denature most proteins [2] [3]. This guide provides a comparative analysis of stability mechanisms between traditional microbial enzymes and extremozymes, offering a structured framework for selecting and engineering biocatalysts tailored to specific application needs.
Fundamental Mechanisms of Enzyme Stability
The stability of an enzyme is governed by a network of intramolecular interactions and structural features that maintain its native, catalytically active conformation. These mechanisms differ significantly between enzymes derived from mesophilic microorganisms and those from extremophiles.
Molecular Determinants of Stability
- Hydrophobic Interactions: In thermophiles, increased hydrophobic core packing reduces water accessibility and enhances thermostability. In contrast, psychrophiles exhibit weaker hydrophobicity to maintain flexibility at low temperatures [2].
- Hydrogen Bonding & Salt Bridges: Thermophilic enzymes contain a higher number of charged residues (Glu, Arg, Lys) that form intricate networks of salt bridges and hydrogen bonds, stabilizing the protein structure at high temperatures [2] [4]. Psychrophilic enzymes minimize these interactions to reduce rigidity [2].
- Amino Acid Composition: Biases in amino acid usage significantly influence stability. Thermophiles show preferences for aromatic and charged residues, while psychrophiles increase glycine content to enhance backbone flexibility [2].
- Structural Compactness: Thermophilic proteins often feature shorter surface loops, reduced surface-to-volume ratios, and increased secondary structure stabilization [2]. Piezophiles (pressure-adapted) incorporate larger volume cavities in their structures to accommodate compression [4].
Membrane and Cofactor Adaptations
Beyond the protein itself, stability is influenced by cellular components. Thermophiles utilize ether-linked lipids in their membranes for enhanced heat resistance, while psychrophiles increase unsaturated fatty acids to maintain membrane fluidity in cold environments [2] [3]. Some alkaliphiles employ specialized cytochrome c and surface proteins to maintain proton homeostasis crucial for function at high pH [4].
Comparative Analysis: Microbial vs. Extremophile Enzymes
The table below summarizes key stability characteristics and molecular adaptations of enzymes from different biological sources.
Table 1: Comparative Stability Profiles of Microbial and Extremophile Enzymes
| Enzyme Source | Optimal Temp Range | Structural Features | Molecular Adaptations | Industrial Applications |
|---|---|---|---|---|
| Psychrophiles (Cold-adapted) | <15-20°C [4] | Heat-labile, flexible active sites, less compact structures [2] | Weaker hydrophobicity, fewer salt bridges/H-bonds, more glycine, unsaturated membrane lipids [2] [3] | Food processing, environmental bioremediation, biotechnology [3] |
| Mesophiles (Traditional microbes) | 20-45°C [2] | Moderate structural rigidity, standard active site accessibility | Standard amino acid composition, typical H-bond/salt bridge density | Detergents, baking, brewing, dairy processing [5] |
| Thermophiles (Heat-adapted) | 60->80°C [4] | Compact structures, shortened loops, increased secondary structure [2] | Enhanced hydrophobic packing, additional salt bridges/H-bonds, charged/aromatic residues, ether-linked lipids [2] [4] | PCR (Taq polymerase), starch processing, biofuel production [2] [6] |
| Halophiles (Salt-adapted) | Varies | Highly acidic, negatively charged surfaces [1] | Enriched surface acidic residues (Asp, Glu), solute accumulation for osmotic balance [1] | Biocatalysis in organic media, biosensors, cosmetic formulations [6] [1] |
| Piezophiles (Pressure-adapted) | Varies | Larger volume cavities, pressure-resistant folding [4] | Polar/ hydrophilic amino acid substitutions, specialized chaperones [4] | High-pressure bioreactors, deep-sea biotechnology [3] |
Experimental Protocols for Assessing Enzyme Stability
Standardized experimental protocols are essential for quantitatively comparing enzyme stability across different conditions and enzyme types. Below are key methodologies cited in current research.
Thermostability Assessment
Melting Temperature (Tₘ) Determination
- Objective: To determine the temperature at which 50% of the enzyme population becomes unfolded [7].
- Protocol:
- Prepare purified enzyme solution in appropriate buffer.
- Use differential scanning calorimetry (DSC) or fluorimetry-based thermal shift assays.
- Apply a controlled temperature ramp (e.g., 1°C per minute) while monitoring unfolding.
- Plot the unfolding curve and calculate Tₘ as the inflection point.
- Data Interpretation: Higher Tₘ values indicate greater intrinsic thermostability. The magnitude of thermal shift (ΔTₘ) upon substrate binding can reveal stabilization effects, with smaller enzymes often showing more substantial ΔTₘ [7].
Thermal Inactivation Half-Life
- Objective: To measure the time-dependent loss of activity at a specific temperature.
- Protocol:
- Incubate enzyme samples at constant elevated temperature.
- Withdraw aliquots at regular time intervals and measure residual activity under standard assay conditions.
- Plot log(residual activity) versus time.
- Calculate half-life (t₁/₂) from the inactivation rate constant.
- Data Interpretation: Longer half-lives indicate superior operational stability for industrial processes requiring prolonged catalysis [1].
Stability Under Other Stress Conditions
pH Stability Profile
- Protocol: Incubate enzymes in buffers of varying pH for a fixed duration, then measure residual activity at optimal pH [1].
- Output: pH range retaining >80% activity indicates functional pH breadth.
Solvent Tolerance
- Protocol: Incubate enzymes with various organic solvents (e.g., dimethylformamide, methanol) and measure residual activity after exposure [1].
- Output: Higher residual activity indicates suitability for non-aqueous biocatalysis.
The following diagram illustrates the logical workflow for a comprehensive enzyme stability assessment, integrating the protocols described above.
Diagram 1: Enzyme Stability Assessment Workflow
The Scientist's Toolkit: Key Reagents and Materials
Successful research into enzyme stability requires specific reagents and tools. The following table details essential solutions and their functions in stability experiments.
Table 2: Essential Research Reagent Solutions for Enzyme Stability Studies
| Reagent/Material | Function in Stability Assessment | Example Application Context |
|---|---|---|
| Thermostable Markers (e.g., Taq Polymerase) | Positive controls for high-temperature stability assays; benchmark for engineering efforts [6]. | Validating thermal cycler performance, comparing thermostability of novel isolates. |
| Buffering Systems (e.g., phosphate, Tris, citrate buffers across pH range) | Maintain specific pH conditions during stability challenges; determine pH activity/stability profiles [1]. | Assessing enzyme functionality in acidic (e.g., food) or alkaline (e.g., detergent) processes. |
| Chaotropic Agents (e.g., Urea, Guanidine HCl) | Induce controlled protein unfolding; measure conformational stability via denaturation curves [1]. | Determining the free energy of unfolding (ΔG) and comparing structural rigidity. |
| Organic Solvents (e.g., DMSO, methanol, hexane) | Assess stability and activity in non-aqueous environments relevant to industrial synthesis [1]. | Screening enzymes for biocatalysis in organic media for pharmaceutical intermediate synthesis. |
| Protease Inhibitors (e.g., PMSF, EDTA) | Protect enzymes from proteolytic degradation during purification and storage, preventing misleading stability data [7]. | Maintaining enzyme integrity in crude extracts or during long-term stability studies. |
| Immobilization Supports (e.g., chitosan, silica, epoxy-activated resins) | Enhance operational stability by rigidifying enzyme structure and enabling reuse [1]. | Developing reusable biocatalysts for continuous flow reactors in API manufacturing. |
| Spectrophotometric Assay Kits (e.g., based on pNA, CDNB, or other chromogenic substrates) | Enable rapid, high-throughput measurement of residual enzyme activity after stability challenges [1]. | Screening large libraries of enzyme variants for improved stability in directed evolution campaigns. |
Emerging Strategies for Enhancing Enzyme Stability
Enzyme Engineering and Miniaturization
Rational design and directed evolution represent powerful approaches for improving enzyme stability. Rational design focuses on introducing specific mutations to enhance stabilizing interactions, such as adding salt bridges, disulfide bonds, or optimizing surface charge [1]. Directed evolution mimics natural selection in the laboratory by generating diverse mutant libraries and screening for variants with enhanced stability under desired conditions [1].
A promising strategy is enzyme miniaturization, which involves creating smaller, catalytically active versions of enzymes. Smaller enzymes often demonstrate superior expression yields, faster folding kinetics, reduced misfolding propensity, and potentially enhanced thermostability due to the removal of flexible, non-essential loops that can initiate unfolding [7].
Bioinformatics and AI-Driven Discovery
Advanced computational methods are revolutionizing stability prediction and engineering. Machine learning models trained on protein sequences and structures can now predict mutation effects on stability and function [1]. Tools like EZSpecificity use graph neural networks to accurately predict enzyme-substrate interactions, aiding in the design of enzymes with tailored stability and specificity [8]. Furthermore, metagenomic mining of unculturable extremophiles directly from environmental DNA provides access to a vast reservoir of novel, stable enzymes without the need for laboratory cultivation [3] [6].
Enzyme stability is a multifaceted property dictated by complex molecular interactions. The comparative analysis presented in this guide underscores that extremophile enzymes offer unparalleled stability under extreme conditions, providing valuable blueprints for engineering more robust biocatalysts. However, traditional microbial enzymes remain indispensable for applications under moderate conditions due to their catalytic efficiency and ease of production.
Future advancements will increasingly rely on integrating structural biology insights with AI-driven tools and high-throughput experimental validation. This multidisciplinary approach will accelerate the development of next-generation enzymes with tailor-made stability properties, ultimately driving innovation across industrial and pharmaceutical sectors.
Microbial enzymes are fundamental biocatalysts that accelerate biochemical reactions across numerous industries, including pharmaceuticals, food processing, detergents, and biofuel production [9]. Their commercial success hinges on stability and functional integrity under process-specific conditions. Conventional microbial enzymes, typically sourced from mesophilic microorganisms (growing at 20–45°C), exhibit optimal performance within narrow ranges of temperature, pH, and solvent concentration [10]. Beyond these ranges, they face significant stability challenges that can limit their industrial efficacy.
The growing demand for sustainable bioprocesses has intensified the need for robust enzymes, shifting research focus toward understanding and enhancing the stability profiles of conventional microbial enzymes under stressful industrial conditions [9]. This review systematically compares the stability and limitations of these enzymes, providing a structured analysis of experimental data, stabilization methodologies, and strategic recommendations for industry applications.
Defining Stability and Stress Conditions for Microbial Enzymes
Enzyme stability refers to an enzyme's ability to resist denaturation and maintain its catalytic function when exposed to various deterrents such as extreme pH, organic solvents, high salt concentrations, and thermal stress [11]. Denaturation involves the loss of the enzyme's native secondary, tertiary, or quaternary structure through the application of external stressors, leading to inactivation [11].
For conventional microbial enzymes, common industrial stress conditions include:
- Temperature Stress: Exposure to temperatures significantly higher than their optimal range (typically above 45-50°C) can cause unfolding and aggregation [10].
- pH Stress: Operating in highly acidic (pH < 5) or alkaline (pH > 9) environments can alter charged amino acid residues, disrupting the active site and protein structure [9].
- Chemical Stress: Contact with organic solvents, heavy metals, or denaturing agents can interfere with hydrophobic interactions and hydrogen bonding [12].
- Oxidative Stress: Presence of oxidizing agents can damage susceptible amino acids like cysteine and methionine [13].
Comparative Stability Profiles: Quantitative Analysis
The following tables summarize typical stability profiles of conventional microbial enzymes under various stress conditions, based on experimental data.
Table 1: Thermal Inactivation Parameters of Conventional Microbial Enzymes
| Enzyme Class | Microbial Source | Optimal Temp (°C) | Half-life at 50°C | Inactivation Temp (°C) | Key Stabilizing Factors |
|---|---|---|---|---|---|
| Protease | Bacillus licheniformis | 50-60 | ~30 min | ~65 | Ca²⁺ ions, high substrate concentration |
| Lipase | Pseudomonas alcaligenes | 37-45 | < 60 min | ~55 | Interfacial activation, lid structure |
| α-Amylase | Bacillus amyloliquefaciens | 70 | ~15 min | ~80 | Ca²⁺ ions, chloride ions |
| Cellulase | Trichoderma reesei | 40-50 | ~20 min | ~60 | Glycosylation, substrate binding |
Table 2: Stability of Conventional Enzymes Under pH and Solvent Stress
| Enzyme Class | Optimal pH Range | Activity Retention at pH 5 (%) | Activity Retention at pH 9 (%) | Activity in 20% Organic Solvent (%) |
|---|---|---|---|---|
| Protease (Neutral) | 6.5-7.5 | < 30 | < 40 | < 50 |
| Lipase | 7.0-9.0 | < 20 | > 80 | > 70 |
| α-Amylase | 5.5-6.5 | > 70 | < 25 | < 40 |
| Laccase (Fungal) | 4.0-5.0 | > 80 | < 20 | < 35 |
Molecular Mechanisms of Instability and Denaturation
The stability limitations of conventional microbial enzymes stem from their structural sensitivity to environmental extremes. Under thermal stress, the kinetic energy overwhelms the weak forces—hydrogen bonds, hydrophobic interactions, and van der Waals forces—that maintain the protein's tertiary structure [10]. This leads to unfolding, exposure of hydrophobic regions, and often irreversible aggregation.
Under pH stress, alterations in the ionization state of amino acid side chains disrupt salt bridges and electrostatic networks essential for structural integrity. For instance, acidophiles maintain stability at low pH through a high content of acidic residues on their surface, while conventional enzymes lack such adaptations [14]. In organic solvents, the desolvation of the enzyme molecule and disruption of essential water layers lead to significant rigidity loss and decreased catalytic efficiency [11].
Diagram: Structural Denaturation Pathways of Conventional Enzymes Under Stress
Methodologies for Assessing Enzyme Stability
Experimental Protocols for Stability Characterization
Standardized experimental protocols are essential for generating comparable stability data. The following methodologies are widely employed:
Thermal Stability Assay:
- Prepare enzyme solution in appropriate buffer at optimal pH.
- Aliquot samples into thin-walled PCR tubes.
- Incubate samples at target temperatures (e.g., 40°C, 50°C, 60°C, 70°C) in thermal cyclers or water baths.
- Withdraw aliquots at predetermined time intervals (0, 5, 15, 30, 60 minutes).
- Immediately cool samples on ice and measure residual activity using standard assays.
- Calculate half-life (t₁/₂) and deactivation rate constants from activity decay curves [9].
pH Stability Profile:
- Prepare buffer systems covering pH range 3.0-10.0.
- Incubate enzyme in each buffer for fixed duration (typically 1 hour) at 25°C.
- Measure residual activity under standard conditions.
- Determine optimal pH and stability range from activity profile [11].
Solvent Tolerance Assay:
- Prepare enzyme solutions containing 10-30% (v/v) organic solvents (e.g., methanol, acetonitrile, DMSO).
- Incubate with shaking for 4-24 hours at 25°C.
- Measure residual activity compared to solvent-free control.
- Assess structural integrity via circular dichroism or fluorescence spectroscopy [15].
Diagram: Workflow for Comprehensive Enzyme Stability Assessment
Research Reagent Solutions for Stability Studies
Table 3: Essential Reagents for Enzyme Stability Research
| Reagent/Category | Specific Examples | Function in Stability Assessment |
|---|---|---|
| Buffer Systems | phosphate, Tris-HCl, citrate, glycine | Maintain specific pH during stress tests |
| Thermal Stabilizers | glycerol, sorbitol, Ca²⁺ ions | Protect against thermal denaturation |
| Solvent Additives | polyols, sugars, proline | Counteract solvent-induced denaturation |
| Activity Assay Reagents | chromogenic substrates (pNPP, casein), DNS | Quantify residual enzymatic activity |
| Structural Probes | ANS, Sypro Orange | Detect unfolding and hydrophobic exposure |
| Immobilization Carriers | chitosan, silica nanoparticles, Eupergit C | Provide stabilization via rigid support |
Strategies for Enhancing Enzyme Stability
Immobilization Techniques
Immobilization represents a primary physical method for stabilizing conventional enzymes by restricting their mobility through binding to a solid substrate [11]. This approach significantly enhances resistance to temperature, pH, solvents, and impurities while enabling enzyme reusability and simplified separation from reaction mixtures [11].
The five principal immobilization techniques include:
- Adsorption: Reversible binding via weak forces (salt linkage, hydrogen bonds, ionic bonds) to supports like silica, chitosan, or cellulose [11].
- Covalent Binding: Formation of stable covalent bonds between enzyme functional groups and activated carriers like agarose or Eupergit C [11].
- Encapsulation: Entrapment within polymer matrices or microcapsules [11].
- Cross-linking: Creating intermolecular bonds between enzyme molecules using linkers like glutaraldehyde [11].
- Entrapment: Physical confinement in polymeric networks [11].
Chemical Modification and Engineering Approaches
Protein engineering strategies have demonstrated remarkable success in enhancing enzyme stability:
- Rational Design: Site-directed mutagenesis to target specific amino acid substitutions that enhance stability, requiring detailed 3D structural information [9].
- Directed Evolution: Iterative rounds of random mutagenesis and screening to select variants with improved stability under stress conditions [9].
- Chemical Modification: Covalent attachment of stabilizing polymers (e.g., polyethylene glycol) or carbohydrates to enzyme surfaces [11].
- Additive Incorporation: Use of co-solvents, osmolytes, and compatible solutes to stabilize enzymes in suboptimal environments [15].
The stability profiles of conventional microbial enzymes reveal significant limitations under industrial stress conditions, particularly when compared to extremozymes from thermophiles, psychrophiles, and halophiles [14]. While conventional enzymes typically exhibit half-lives of minutes at elevated temperatures and significant activity loss under pH or solvent stress, extremozymes can maintain functionality for hours or days under similar conditions [14].
This stability gap underscores the importance of strategic enzyme selection based on process requirements. For applications involving moderate conditions, conventional enzymes remain cost-effective solutions. However, for processes requiring extreme temperatures, pH, or solvent tolerance, extremozymes or engineered variants offer superior performance despite potentially higher initial costs [9] [14].
Future research directions should focus on integrating multi-omics approaches with advanced protein engineering to develop next-generation enzymes that combine the catalytic efficiency of conventional enzymes with the robustness of extremozymes [14]. Such advances will significantly expand the application scope of microbial enzymes in sustainable industrial processes.
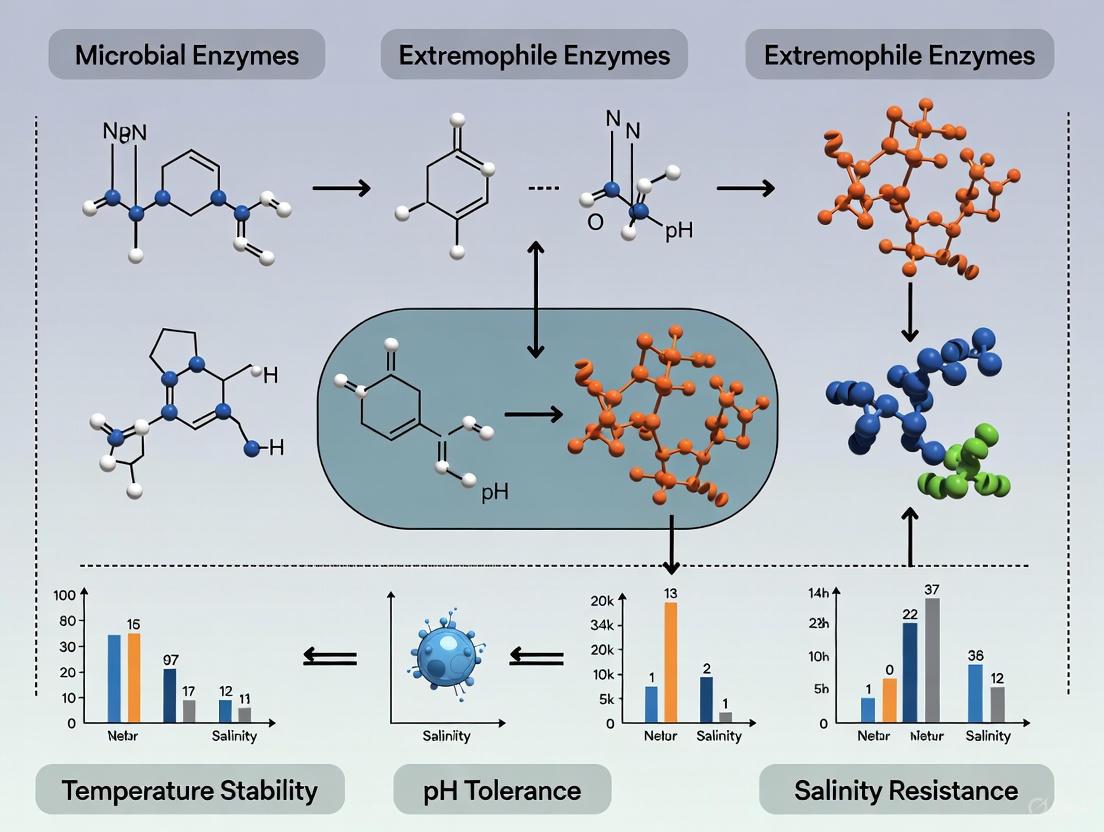
Extremophiles are organisms that thrive in ecological niches characterized by conditions once considered incompatible with life, redefining our understanding of life's resilience and adaptability [16] [6]. These remarkable organisms inhabit environments with extreme temperatures, pH levels, salinity, pressure, and radiation, utilizing unique biochemical adaptations to not only survive but flourish where conventional life would perish [6] [17]. The study of extremophiles has gained significant scientific interest since the discovery of Thermus aquaticus from Yellowstone National Park's hot springs in the 1960s, fundamentally reshaping our perspective on life's boundaries and evolutionary origins [18] [17].
From a comparative enzymology perspective, extremophiles represent a fascinating subject for investigating protein stability and function under physical-chemical extremes. Their enzymes, known as extremozymes, exhibit exceptional stability and catalytic efficiency under conditions that would rapidly denature or inactivate their conventional counterparts from mesophilic organisms [16] [14]. This review systematically compares the adaptive strategies and enzymatic machinery of extremophiles across niche types, with particular emphasis on experimental approaches for quantifying stability parameters that inform both fundamental biology and biotechnological applications in drug development and industrial processes.
Comparative Analysis of Extremophile Niches and Adaptive Strategies
Extremophiles are classified based on the specific environmental parameters of their ecological niches. The table below provides a comprehensive comparison of major extremophile categories, their niche characteristics, and corresponding adaptive mechanisms at cellular and molecular levels.
Table 1: Extremophile Classification, Niche Characteristics, and Adaptive Strategies
| Extremophile Type | Niche Characteristics | Representative Species | Key Adaptive Strategies |
|---|---|---|---|
| Thermophiles | Temperatures exceeding 40°C up to 122°C [16] [18] | Methanopyrus kandleri (122°C) [18] | Thermostable enzymes with reinforced salt bridges, hydrophobic interactions, hydrogen bonding [17] |
| Psychrophiles | Temperatures below -17°C [16] | Psychromonas ingrahamii (-12°C) [18] | Cold-adapted enzymes with structural flexibility; antifreeze proteins [16] [17] |
| Acidophiles | pH below 5 [16] | Picrophilus oshimae (pH 0) [18] | Reinforced membrane composition; proton export machinery [18] |
| Alkaliphiles | pH above 9 [16] | Bacillus alkaliphilus (pH 9.5-10.5) [18] | Specialized membrane transporters; cytoplasmic acidification mechanisms |
| Halophiles | Salt concentrations >3.5% [16] | Halorhodospira halophila (13% NaCl) [18] | Compatible solute synthesis; ion homeostasis; specialized membrane composition [17] |
| Piezophiles | High-pressure environments [16] | Moritella yayanosii (100 MPa) [18] | Piezolyte production; membrane fluidity modulation via unsaturated fatty acids [17] |
| Radiophiles | Radiation levels exceeding natural background [16] | Deinococcus radiodurans [6] | Efficient DNA repair mechanisms; oxidative damage protection proteins [6] [17] |
Many extremophiles are polyextremophiles, adapted to multiple simultaneous stresses [14] [17]. For example, Saccharolobus solfataricus (formerly Sulfolobus solfataricus) thrives at 80°C and pH 2.0-4.0, requiring adaptations for both high temperature and extreme acidity [18]. These complex niche requirements drive the evolution of sophisticated, integrated adaptation mechanisms that confer stability across several physicochemical parameters simultaneously.
Experimental Methodologies for Assessing Enzyme Stability
The comparative analysis of enzyme stability between conventional microbial enzymes and extremozymes requires specialized experimental protocols designed to measure functional integrity under extreme conditions. Below are detailed methodologies for key stability assays cited in extremophile enzyme research.
Thermal Stability Profiling
Objective: Quantify enzyme thermostability by measuring residual activity after exposure to elevated temperatures. Protocol:
- Prepare enzyme solution in appropriate buffer at pH optimum
- Aliquot samples into thin-walled PCR tubes
- Perform heat challenge in thermal cycler: incubate at temperature range from 50°C to 100°C for time intervals (15-120 minutes)
- Immediately cool samples on ice for 10 minutes
- Measure residual enzymatic activity using standard assay conditions
- Calculate half-life (t½) at each temperature from activity decay curves Applications: Particularly essential for characterizing thermozymes from organisms like Pyrococcus furiosus (optimal growth at 100°C) [18] and comparing them to mesophilic counterparts.
pH Tolerance Assessment
Objective: Determine enzymatic activity and stability across pH spectrum. Protocol:
- Prepare buffer systems covering pH range 1.0-12.0 (e.g., glycine-HCl, citrate, phosphate, Tris, glycine-NaOH)
- Incubate enzyme in each buffer for 1 hour at 4°C
- Measure initial activity under respective pH conditions
- For stability assessment, return aliquots to optimal pH and measure recovered activity
- Plot activity vs. pH to determine optimal range and stability profile Applications: Critical for characterizing acidophiles (e.g., Picrophilus oshimae, pH 0) and alkaliphiles (e.g., Bacillus alkaliphilus, pH 10.5) [18].
Halostability Measurement
Objective: Quantify enzyme function and stability at high salt concentrations. Protocol:
- Prepare reaction mixtures with NaCl concentrations ranging from 0-5 M
- Measure enzyme activity at each salt concentration
- For stability assessment, pre-incubate enzyme in different NaCl concentrations for 24 hours
- Measure residual activity under optimal assay conditions
- Compare kinetics parameters (Km, Vmax) at various salt concentrations Applications: Essential for studying halophiles like Halorhodospira halophila which thrives at 13% NaCl [18].
The following diagram illustrates the experimental workflow for systematic characterization of extremophile enzyme stability:
Quantitative Comparison of Enzyme Stability Parameters
The exceptional utility of extremozymes in biotechnology and industrial processes stems from their quantified stability advantages over conventional enzymes. The table below summarizes comparative experimental data highlighting these differences across stability parameters.
Table 2: Comparative Stability Parameters of Microbial vs. Extremophile Enzymes
| Enzyme Type | Source Organism | Optimal Activity Temperature | Thermal Half-life (at 90°C) | pH Stability Range | Salt Tolerance |
|---|---|---|---|---|---|
| Conventional Protease | Bacillus subtilis (mesophile) | 37-45°C | <5 minutes | 5.5-8.5 | Low (<0.5 M NaCl) |
| Thermostable Protease | Thermus aquaticus | 70-80°C | >60 minutes [17] | 6.0-9.0 | Moderate (1-2 M NaCl) |
| Thermostable Polymerase | Pyrococcus furiosus | 100°C [18] | >4 hours [18] | 6.5-8.0 | Low-Moderate |
| Cold-adapted Lipase | Psychromonas ingrahamii | 4-15°C [18] | N/A (thermolabile) | 7.0-9.0 | High (2-3 M NaCl) [14] |
| Halotolerant Esterase | Halorhodospira halophila | 37-45°C | <30 minutes | 7.5-10.5 | Extreme (4-5 M NaCl) [18] |
| Acidophilic Amylase | Saccharolobus solfataricus | 80°C [18] | >90 minutes | 2.0-4.0 [18] | Low (<0.5 M NaCl) |
Structural analysis reveals the molecular basis for these stability differences. Thermozymes exhibit strengthened hydrophobic cores, increased salt bridge networks, and enhanced hydrogen bonding compared to their mesophilic counterparts [17]. Psychrophilic enzymes achieve cold activity through reduced proline/arginine content, fewer salt bridges, and increased surface hydrophilicity that provides structural flexibility at low temperatures [14]. Halophilic enzymes display high surface acidity with abundant aspartic and glutamic acid residues that coordinate hydrated salt ions, maintaining solvation and function at near-saturating salt concentrations [17].
The Scientist's Toolkit: Essential Research Reagents
Research into extremophile enzymes requires specialized reagents and materials designed to maintain enzyme integrity and facilitate activity measurements under extreme conditions. The following table details key solutions and their applications in extremophile enzymology.
Table 3: Essential Research Reagents for Extremophile Enzyme Characterization
| Reagent/Material | Function | Application Example |
|---|---|---|
| Thermostable Enzyme Substrates (e.g., p-nitroaniline-linked peptides) | Colorimetric activity detection at high temperatures | Measuring protease activity in thermophiles like Pyrococcus furiosus at 100°C [18] |
| Specialized Buffer Systems (e.g., HEPES, MES, CHES) | Maintain precise pH under extreme conditions | pH stability profiling across range 1.0-12.0 for acidophiles/alkaliphiles |
| Osmoprotectants (e.g., betaine, trehalose) | Stabilize enzymes during freezing/thawing | Preserving activity in psychrophilic enzyme preparations |
| Halostability Buffers (with 0-5 M NaCl/KCl) | Maintain ionic strength for halophile function | Characterizing salt-dependent activity in halophiles like Halorhodospira halophila [18] |
| Chaotrope Resistance Agents (e.g., sorbitol, glycerol) | Protect against chemical denaturants | Assessing enzyme stability in industrial process conditions |
| Piezophilic Culture Chambers | Maintain high-pressure conditions | Studying enzymes from piezophiles like Moritella yayanosii (100 MPa) [18] |
The following diagram illustrates the logical relationship between extremophile niches, their enzyme adaptations, and resulting biotechnological applications:
The systematic comparison of extremophiles and their enzymes reveals a remarkable continuum of biological adaptation to environmental extremes. From thermal springs approaching 122°C to hypersaline lakes and acidic geothermal fields, these organisms have evolved enzymatic machinery with precisely calibrated stability parameters that enable life at the physical-chemical limits. The quantitative data and experimental methodologies presented herein provide researchers with standardized approaches for comparative enzymology in extreme conditions.
For drug development professionals, extremozymes offer more than just process advantages; they represent sources of novel catalytic mechanisms and structural scaffolds with potential therapeutic applications. The continued exploration of Earth's most challenging environments through metagenomic and single-cell genomic approaches promises to reveal even more extraordinary enzymatic diversity with applications spanning medicine, biotechnology, and industrial catalysis [6] [18]. As climate change alters global ecosystems, understanding these limits of life becomes increasingly crucial for developing resilient biotechnological solutions in a rapidly changing world.
The study of extremophiles—organisms that thrive in conditions once deemed incompatible with life—has fundamentally reshaped our understanding of protein stability and function. Among these remarkable organisms, thermophiles and psychrophiles represent two evolutionary extremes, inhabiting environments characterized by persistently high or low temperatures, respectively. Their enzymes, known as extremozymes, have undergone specialized adaptations to maintain activity and structural integrity under these challenging conditions. For thermophiles, the imperative is to resist denaturation and unfolding at high temperatures, achieved through enhanced structural rigidity. In contrast, psychrophiles face the opposite challenge: maintaining catalytic flexibility and dynamics at temperatures that would typically render mesophilic proteins inert. This guide provides a detailed, evidence-based comparison of the molecular strategies employed by these extremophiles, framing the discussion within broader research on comparative enzyme stability and providing actionable experimental data and protocols for researchers in biotechnology and drug development.
Structural & Molecular Basis of Adaptation
The distinct thermal challenges faced by thermophiles and psychrophiles have driven the evolution of unique structural solutions at the molecular level. The table below summarizes the key adaptive traits observed in proteins from these organisms.
Table 1: Molecular Adaptations in Thermophilic and Psychrophilic Proteins
| Adaptive Trait | Thermophilic Proteins | Psychrophilic Proteins |
|---|---|---|
| Amino Acid Propensity | Increased Arg, Tyr; Decreased Cys, Ser [19] [20] | More small residues (e.g., Gly); Reduced bulky hydrophobic cores [21] |
| Secondary Structure | Higher fraction of residues in α-helices; Avoids Pro in helices [19] | Reduced helical content; Longer surface loops [21] |
| Stabilizing Interactions | Increased salt bridges & side-chain H-bonds; Dense ionic networks [22] [19] | Fewer salt bridges & H-bonds; Weakened intramolecular interactions [21] |
| Hydrophobic Core | Increased packing density; More aromatic clusters [22] [21] | Reduced packing density; Fewer aromatic clusters [21] |
| Overall Structural Rigidity | High; reinforced by networks of charges [22] | Low; maintains flexibility at low temperatures [23] [21] |
These molecular differences are not random but form a coherent adaptive strategy. Thermophilic proteins enhance rigidity through a multi-faceted approach: charged residues like arginine and glutamate form intricate networks of salt bridges that cross-link different structural elements, while a compact hydrophobic core and more extensive hydrogen bonding provide internal stability [22] [19]. Psychrophiles, conversely, employ a strategy of structural loosening. They reduce the number and strength of stabilizing interactions, possess a less compact core, and incorporate more small residues like glycine, which collectively increase backbone flexibility and allow functional motion at low energy costs [21].
Visualizing the Key Structural Differences
The following diagram synthesizes the core structural concepts from the research, illustrating the network-based stabilization in thermophiles versus the reduced interaction strategy in psychrophiles.
Experimental Data & Quantitative Comparisons
Empirical data is crucial for understanding the functional outcomes of these structural adaptations. The following table quantifies key biophysical and functional parameters, drawing from direct experimental measurements and analyses.
Table 2: Experimental Data on Protein Stability and Dynamics
| Parameter | Thermophiles | Psychrophiles | Experimental Method |
|---|---|---|---|
| Melting Temperature (Tm) | Can exceed 100°C [23] | Can be 20-30°C below mesophiles [23] | Differential Scanning Calorimetry (DSC) |
| Optimal Activity Range | Often 70°C - 120°C [21] | Often < 20°C [21] | Enzyme kinetics assay |
| Protein Diffusion Coefficient (DG) | Lower at respective physiological temps (e.g., ~1.5 Ų/ns for A. aeolicus at 370K) [23] | Higher at respective physiological temps (e.g., ~1.8 Ų/ns for P. arcticus at 317K) [23] | Quasi-elastic Neutron Scattering (QENS) |
| Dynamic Arrest Temperature | Coincides with cell death temperature [23] | Decoupled from cell death; occurs ~22°C above [23] | QENS & Molecular Dynamics (MD) |
| Catalytic Activity (kcat) | Often lower at mesophilic temps | Significantly higher at low temps (0-10°C) | Spectrophotometric assays |
A critical finding from recent research is the different relationship between protein dynamics and cell viability in these extremophiles. In thermophiles like Aquifex aeolicus, a sharp slowdown in protein diffusion—a dynamic arrest caused by the unfolding of a small fraction of the proteome—occurs precisely at the organism's cell death temperature (~370K) [23]. This suggests that the loss of proteome solubility and dynamics is a key factor in thermal death for these organisms. In stark contrast, the psychrophile Psychrobacter arcticus experiences this same dynamic arrest at a temperature (~317K) far above its cell death temperature (~295K) [23]. This indicates that psychrophiles do not die from a gelling of their cytoplasm, but likely from the loss of specific, critical enzymatic functions at low temperatures, underscoring a fundamental decoupling between proteome stability and viability.
Detailed Experimental Protocols
To facilitate reproducibility and further research, this section outlines key methodologies used to generate the comparative data discussed in this guide.
Protocol 1: X-Ray Crystallography for Structural Determination
This protocol is used to solve high-resolution structures of extremozymes, enabling atomic-level analysis of stabilizing features like salt bridges and hydrophobic packing [24].
- 1. Protein Expression and Purification: Clone the target gene into an appropriate expression vector (e.g., for E. coli). Express the protein and purify it using chromatography methods (e.g., affinity, size exclusion) to homogeneity.
- 2. Crystallization: Screen for crystallization conditions using robotic systems and commercial screens. Optimize hits to grow large, single crystals suitable for diffraction.
- 3. Data Collection: Flash-cool the crystal in liquid nitrogen. Collect X-ray diffraction data at a synchrotron source. For example, the structure of a thermophilic malate dehydrogenase (1GV1) was determined at 2.50 Å resolution [24].
- 4. Structure Solution and Refinement: Solve the phase problem using molecular replacement or other phasing methods. Iteratively refine the model using programs like CNS, building the structure and optimizing its fit to the electron density map. Key refinement statistics to report include R-value Work and R-value Free [24].
Protocol 2: Quasi-Elastic Neutron Scattering (QENS) for Protein Dynamics
QENS measures the diffusive motion of proteins within the crowded cellular environment, providing insights into cytoplasmic fluidity and rigidity [23].
- 1. Sample Preparation: Grow bacterial cells (e.g., P. arcticus, A. aeolicus) to mid-log phase. To highlight the signal from proteins, the cells are typically transferred into a D₂O-based buffer, as deuterium reduces the neutron scattering signal from the solvent.
- 2. Data Acquisition: Load the sample into a sealed cell compatible with the neutron spectrometer. Experiments are performed on instruments like the IN16B spectrometer at the Institute Laue Langevin. Data is collected over a defined temperature range (e.g., from 200K to 370K) that encompasses the organism's physiological and lethal temperatures.
- 3. Data Analysis: The obtained spectra, the incoherent dynamic structure factor S(Q,E), are fitted with a model that decomposes the dynamics into global diffusion (translational and rotational motion of the whole protein) and local diffusion (internal motions). The global diffusion coefficient (DG) is a key output parameter, revealing how freely proteins move in the cytoplasm [23].
Workflow for Comparative Stability Analysis
The following diagram outlines a logical workflow for a research project aimed at comparing the stability of extremophile enzymes, integrating the protocols described above.
The Scientist's Toolkit: Key Research Reagents & Solutions
This section catalogs essential materials and tools derived from the search results that are pivotal for research in this field.
Table 3: Essential Research Reagents and Solutions
| Reagent / Solution | Function / Application | Example / Key Feature |
|---|---|---|
| Taq DNA Polymerase | High-temperature DNA amplification (PCR) [25] [6] | Derived from Thermus aquaticus; Thermostable [25] |
| Halomonas bluephagenesis | Halophilic chassis for open, non-sterile fermentation [25] | Engineered to produce bioplastics (PHA) [25] |
| CRISPR-Cas Systems | Gene editing in extremophile chassis [25] [6] | Enables precise genomic modifications in non-model organisms [25] |
| Porousin Promoter Library | Regulating gene expression in halophiles [25] | A constitutive promoter library for Halomonas [25] |
| Specialized Growth Media | Cultivation of extremophiles under specific conditions [21] | High-salt for halophiles; specific pH for acidophiles/alkaliphiles [26] |
| L-Asparaginase | Food processing & cancer treatment [6] | Sourced from halotolerant Bacillus strains [6] |
The comparative analysis of thermophilic and psychrophilic proteins reveals two elegant, opposing solutions to the problem of environmental extreme: the reinforcement of structural rigidity versus the optimization of catalytic flexibility. These molecular blueprints, characterized by distinct amino acid usage, stabilizing interaction networks, and dynamic properties, are not merely academic curiosities. They provide a rich repository of design rules and components for biotechnology. Understanding these principles enables researchers to engineer enzymes with tailored stability, develop robust microbial chassis for industrial processes, and explore novel mechanisms for drug development where enzyme stability or flexibility is a critical factor. The continued integration of structural biology, biophysical dynamics, and synthetic biology will undoubtedly unlock further transformative applications inspired by life at the edge.
Genomic and Metabolic Adaptations Supporting Enzyme Function in Extreme Conditions
The study of extremophilic organisms—those thriving in conditions once considered inhospitable to life—has fundamentally reshaped our understanding of biological adaptability. Thermophiles and psychrophiles, adapted to high (55-80°C) and low (<15°C) temperatures respectively, represent remarkable case studies in evolutionary adaptation [27] [14]. These organisms possess specialized enzymatic machinery, known as extremozymes, that catalyze chemical reactions under conditions that would denature or inactivate their mesophilic counterparts [14]. The genomic and metabolic adaptations underlying these capabilities are not merely academic curiosities; they offer blueprints for biotechnological innovation, with applications spanning pharmaceutical development, industrial biocatalysis, and sustainable manufacturing [14] [28]. This review systematically compares the genomic foundations and metabolic network properties of extremophiles against mesophilic reference points, providing researchers with a structured framework for understanding enzyme stability-activity tradeoffs across the thermal spectrum.
Genomic Blueprints of Thermal Adaptation
Genome Architecture and Composition
Comparative genomic analyses reveal distinct evolutionary strategies in thermophiles and psychrophiles. Psychrophiles generally possess significantly larger genomes and more coding sequences (CDS) than thermophiles, suggesting a genetic repertoire for environmental sensing and metabolic flexibility in cold environments [27]. Conversely, thermophiles exhibit relatively compact genomes with limited variation in size, reflecting selective pressures for efficiency and stability at elevated temperatures [27].
A fundamental distinction lies in nucleotide composition. Thermophiles consistently demonstrate higher genomic G+C content, particularly at the first codon position, which enhances DNA stability through additional hydrogen bonding [27]. Psychrophiles, by contrast, favor A+T-rich codons, which reduce energy requirements for transcription and translation while potentially maintaining DNA flexibility at low temperatures [27].
Table 1: Comparative Genomic Features of Temperature-Adapted Microorganisms
| Genomic Feature | Thermophiles | Psychrophiles | Mesophiles |
|---|---|---|---|
| Average Genome Size | Smaller | Significantly Larger | Highly Variable |
| Number of Genes (CDS) | Fewer | Significantly More | Variable |
| Genomic G+C Content | Higher | Lower | Intermediate |
| Codon Preference | G+C-rich (GGC, GCG, GCC) | A+T-rich (TTA, AAA, ATT) | Balanced |
| Amino Acid Enrichment | Tyrosine (Y), Glutamate (E), Leucine (L) | Threonine (T), Methionine (M), Phenylalanine (F) | Mixed |
The distinct thermal niches of extremophiles are reflected in their proteomic compositions. Nearly two-thirds (13/20) of amino acids show significantly different abundance patterns when comparing extremophiles to mesophiles [27]. Thermophiles exhibit significant enrichment of tyrosine (Y), glutamate (E), and leucine (L), which contribute to increased hydrophobic interactions and salt bridges that stabilize protein structures at high temperatures [27]. Psychrophiles display increased abundance of threonine (T), methionine (M), phenylalanine (F), serine (S), and tyrosine (Y), which promote molecular flexibility and maintain catalytic activity at low temperatures [27] [14].
These compositional differences directly impact enzyme structure-function relationships. Psychrophilic enzymes often feature reced hydrogen bonding, fewer salt bridges, and reduced use of proline and arginine in loop regions, all contributing to enhanced molecular flexibility in cold environments [14]. Thermophilic enzymes counter thermal denaturation through increased hydrophobic interactions, additional disulfide bonds, and more compact oligomeric states [14].
Metabolic Network Adaptations
Metabolic Network Architecture
Genome-scale metabolic modeling reveals distinctive network properties adapted to extreme thermal environments. Psychrophiles, despite their larger genomes, maintain the lowest number of exchange reactions (for nutrient transport and waste excretion), significantly fewer than mesophiles [27]. This suggests reduced environmental interaction or more autonomous metabolic networks in cold-adapted organisms.
Both thermophiles and psychrophiles exhibit significantly fewer exchange reactions compared to mesophiles, indicating that extremophiles generally have more constrained interactions with their external environment compared to mesophilic organisms [27]. This metabolic "self-sufficiency" may represent a fundamental adaptation to environments where nutrient availability is limited or unpredictable.
Table 2: Metabolic Network Properties Across Thermal Adaptation Classes
| Metabolic Property | Thermophiles | Psychrophiles | Mesophiles |
|---|---|---|---|
| Number of Metabolic Reactions | Lower | Highest | Intermediate |
| Exchange Reactions | Significantly Reduced | Lowest | Highest |
| Nutrient Import | Intermediate | Significantly Reduced | High |
| Metabolite Export | Significantly Reduced | Intermediate | High |
| Predicted Growth Rates | Lower | Higher | Variable |
Specialized Metabolic Pathways and Biochemical Reactions
Extremophiles employ specialized metabolic strategies tailored to their thermal niches. Psychrophiles maintain higher growth rates in their native environments, supported by metabolic networks that facilitate rapid nutrient assimilation and energy generation despite thermodynamic constraints [27]. Both extremophile groups exhibit distinct active metabolic reactions enriched with unique biochemical processes essential for coping with environmental stressors [27].
At the protein level, psychrophiles produce ice-binding proteins (IBPs) including antifreeze proteins that lower the freezing point of body fluids and prevent ice crystal formation [28]. Thermophiles invest in elaborate chaperone systems and heat shock proteins that prevent protein aggregation and facilitate refolding under thermal stress [28]. These specialized adaptations represent significant metabolic investments that are reflected in their respective network architectures.
Experimental Approaches for Investigating Extremozyme Adaptations
Enzyme Proximity Sequencing (EP-Seq)
Enzyme Proximity Sequencing (EP-Seq) represents a cutting-edge deep mutational scanning method that simultaneously resolves both stability and activity phenotypes for thousands of enzyme variants [29]. This approach leverages peroxidase-mediated radical labeling with single-cell fidelity to dissect the effects of mutations on folding stability and catalytic activity in a single experiment.
The core workflow involves two parallel branches:
- Expression Analysis: Enzyme variants are displayed on yeast surfaces, stained with fluorescent antibodies, and sorted via FACS into bins based on expression levels, which serve as a proxy for folding stability.
- Activity Analysis: A horseradish peroxidase-mediated phenoxyl radical coupling reaction converts oxidase activity into a fluorescent label on the cell wall, enabling sorting based on catalytic function.
Following sorting, next-generation sequencing of both branches enables computational determination of fitness scores for both expression (stability) and activity for each variant [29]. This powerful approach has revealed fundamental biophysical principles, including activity-based constraints that limit folding stability during natural evolution and identified distant "hotspot" regions where mutations can improve catalytic activity without sacrificing stability [29].
Multi-Omics Integration for Extremophile Characterization
Advanced multi-omics approaches provide comprehensive insights into extremophile adaptation by integrating complementary data layers [14] [28]. A representative workflow includes:
- Metagenomic Sequencing: 16S rRNA marker gene analysis or shotgun metagenomics for community profiling and functional gene identification.
- Metatranscriptomics: RNA-Seq analysis to identify actively expressed genes and regulatory responses to environmental conditions.
- Metaproteomics: Mass spectrometry-based protein identification and quantification to validate enzymatic expression and post-translational modifications.
This integrated approach was successfully applied to Thermus filiformis, revealing temperature-dependent physiological changes and identifying numerous thermostable enzymes with biotechnological potential, including amylases, pyrophosphatases, glucosidases, and galactosidases [14]. Such multi-omics frameworks enable researchers to move beyond gene catalogs to understand functional enzymatic adaptations in extreme environments.
The Scientist's Toolkit: Essential Research Reagents and Platforms
Table 3: Key Research Reagents and Platforms for Extremophile Enzyme Studies
| Reagent/Platform | Function | Application Example |
|---|---|---|
| ModelSEED Platform | Genome-scale metabolic network reconstruction | Creating metabolic models from genomic data [27] |
| Yeast Surface Display | Protein expression and stability screening | Displaying enzyme variants for EP-Seq [29] |
| Horseradish Peroxidase (HRP) | Proximity labeling catalyst | Converting enzyme activity to fluorescent signal [29] |
| Tyramide-488 | Fluorescent labeling reagent | Activity-dependent cell staining in EP-Seq [29] |
| Illumina NovaSeq 6000 | High-throughput sequencing | Variant identification in DMS studies [29] |
| FACS Systems | Cell sorting based on fluorescence | Separating variants by expression/activity [29] |
| Site Saturation Mutagenesis | Comprehensive variant library generation | Creating mutant libraries for DMS [29] |
The comparative analysis of genomic and metabolic adaptations in extremophiles reveals fundamental design principles for enzyme function under thermal stress. Thermophiles achieve stability through G+C-rich genomes, compact metabolic networks, and amino acid compositions that favor rigid protein cores, while psychrophiles leverage A+T-rich codons, extensive genetic repertoires, and flexible enzyme architectures for cold activity. These contrasting strategies illustrate the evolutionary trade-offs between stability and activity that constrain natural enzyme evolution.
For pharmaceutical and industrial applications, these insights enable rational engineering of biocatalysts with tailored stability-activity profiles. The experimental frameworks outlined—particularly EP-Seq and multi-omics integration—provide roadmap methodologies for systematic enzyme characterization and engineering. As extremophile research continues to unveil nature's solutions to environmental challenges, the translation of these principles to biotechnology promises more robust, efficient, and specialized enzymes for drug development, biocatalysis, and sustainable manufacturing.
From Discovery to Application: Harnessing Extremozymes in Biomedicine and Industry
The pursuit of novel enzymes, particularly those from extremophiles (extremozymes), is a critical frontier in biotechnology and drug development [30] [18]. These enzymes function in conditions that are inhospitable to most life, and their stability offers immense potential for industrial catalysis, including in pharmaceutical synthesis [14] [30]. Accessing this resource, however, hinges on advanced discovery techniques. For decades, culture-based methods were the cornerstone of microbiology. More recently, metagenomic mining has emerged as a powerful, culture-independent approach [31] [32]. This guide provides an objective comparison of these two paradigms, framing them within ongoing research on the comparative stability of microbial versus extremophile enzymes. We summarize experimental data, detail protocols, and provide key resources to inform the strategies of researchers and scientists.
Principle and Workflow Comparison
The fundamental distinction between these techniques lies in their requirement for microbial cultivation.
- Culture-Based Methods rely on isolating and growing microorganisms in the laboratory on specific nutrient media. Once isolated, enzymes can be purified and characterized from these cultures [32].
- Metagenomic Mining bypasses the cultivation step entirely. It involves directly extracting environmental DNA (eDNA) from a sample, sequencing it, and using computational tools to identify and predict the functions of enzyme-encoding genes [31] [32].
The following diagram illustrates the core workflows for each discovery pipeline.
Diagram: Enzyme Discovery Workflows
Performance and Application Analysis
The choice between culture-based and metagenomic methods involves trade-offs in scope, sensitivity, and functional output. The table below summarizes their comparative performance based on published studies.
Table 1: Comparative Analysis of Culture-Based and Metagenomic Discovery Techniques
| Feature | Culture-Based Methods | Metagenomic Mining |
|---|---|---|
| Fundamental Principle | Isolation and growth of microorganisms in the lab [32]. | Direct extraction and analysis of DNA from environmental samples [31] [32]. |
| Theoretical Coverage | Targets only the culturable minority (~1% of microbes) [30] [32]. | Provides access to the entire microbial community, including the uncultured majority [31] [32]. |
| Functional Output | Provides direct evidence of enzyme activity and allows for experimental characterization of stability and function [32]. | Predicts function from genetic sequence; requires heterologous expression for experimental validation [31]. |
| Key Limitations | Misses vast microbial diversity; labor-intensive and slow [33] [32]. | Does not prove enzyme function or stability directly; high computational cost; gene expression challenges [31] [32]. |
| Pathogen Detection (Sensitivity) | Gold standard for confirming viable pathogens but can be slow (e.g., 15-28 hours for positive culture) [34]. | High sensitivity; can detect pathogens at abundances as low as 0.01% and is less affected by prior antibiotic use [34] [35]. |
| Best Applications | Functional validation, physiological studies, and obtaining stable enzyme-producing strains for industrial scale-up [32] [36]. | Biodiversity exploration, discovery of novel gene sequences from uncultured hosts, and rapid pathogen screening [31] [34]. |
Detailed Experimental Protocols
To ensure reproducibility, this section outlines standard protocols for both techniques as applied in recent research.
Culture-Based Protocol for Pathogen Isolation
This protocol is adapted from a study detecting foodborne pathogens in Malaysian produce [33].
- Sample Homogenization: A 10 g sample is added to 270 mL of a 0.85% saline solution in a stomacher bag and homogenized for 2 minutes.
- Selective Enrichment: The homogenate is added to specific enrichment broths (e.g., Buffered Peptone Water for Salmonella) and incubated under optimal conditions for the target pathogen (e.g., 37°C for 24 hours).
- Plating and Isolation: The enriched broth is serially diluted in a 0.85% saline solution. The dilutions are plated onto selective agar media (e.g., XLD agar for Salmonella) using the spread plate method and incubated.
- Colony PCR Confirmation: Presumptive positive colonies are picked, and DNA is extracted via a boiled-cell method (heating at 100°C for 10 min). Conventional PCR with species-specific primers is used for final confirmation.
Shotgun Metagenomics Protocol for Microbiome Analysis
This protocol is synthesized from studies on oil reservoir microbiota and foodborne pathogens [33] [36].
- Genomic DNA (gDNA) Extraction: gDNA is extracted directly from the environmental or clinical sample using commercial kits (e.g., Nucleospin Food Kit). The extraction includes steps to lyse cells and purify the DNA, often with modifications to improve yield from complex matrices.
- Library Preparation and Sequencing: The extracted gDNA is fragmented, and adapters are ligated to create a sequencing library. The library is then subjected to high-throughput shotgun sequencing on a platform such as Illumina.
- Bioinformatic Analysis:
- Quality Control & Assembly: Raw sequence reads are quality-filtered and trimmed. High-quality reads are assembled de novo into longer contigs.
- Binning & Taxonomic Profiling: Contigs are binned into Metagenome-Assembled Genomes (MAGs) based on sequence composition and abundance. Taxonomic classifiers like Kraken2/Bracken are used to determine microbial community composition [35].
- Functional Annotation: Genes are predicted within the contigs and MAGs. These are annotated against databases (e.g., KEGG, NCBI-NR) to identify putative enzymes, antibiotic resistance genes (ARGs), and virulence factors.
Case Study: Integrated Analysis of Petroleum Reservoirs
A study on high-salinity petroleum reservoirs in Tatarstan demonstrates the power of combining both methods [36].
- Metagenomic Analysis: 16S rRNA sequencing and reconstruction of 75 MAGs revealed a community dominated by sulfidogenic bacteria (e.g., Desulfobacterota) and fermentative bacteria (e.g., Halanaerobiaeota), along with methanogenic archaea (Methanohalophilus).
- Culture-Based Validation: Researchers isolated 20 pure cultures, including genera like Halanaerobium and Geotoga.
- Integrated Finding: The isolated fermentative bacteria were confirmed to produce oil-displacing metabolites (acids, alcohols, gases), highlighting their biotechnological potential for microbial enhanced oil recovery (MEOR). However, metagenomic data also showed that these metabolites could stimulate sulfidogenic bacteria, which cause corrosion. This integrated approach was crucial for designing an MEOR strategy that enhances recovery while suppressing corrosive side effects.
The Scientist's Toolkit: Key Research Reagents
Table 2: Essential Reagents and Kits for Enzyme Discovery Research
| Item | Function/Application | Example Use Case |
|---|---|---|
| Selective Agar Media | Supports the growth of specific microbial taxa while inhibiting others. | Isolating Listeria spp. from food samples using selective agars [33]. |
| Nucleospin Food Kit | Extracts high-quality genomic DNA from complex biological samples like food. | Preparing DNA for shotgun metagenomic sequencing of foodborne pathogens [33]. |
| Kraken2/Bracken | A system for fast, sensitive taxonomic classification of metagenomic sequences and abundance estimation. | Identifying and quantifying pathogens in simulated food metagenomes down to 0.01% abundance [35]. |
| Heterologous Expression Hosts | Surrogate organisms (e.g., E. coli) used to express and produce proteins from cloned foreign genes. | Producing and characterizing a novel esterase discovered from a compost metagenome [31]. |
Both culture-based and metagenomic techniques are indispensable for modern enzyme discovery. Culture-based methods provide the undeniable advantage of direct functional validation and access to viable isolates, which remain crucial for understanding physiology and scaling production. In contrast, metagenomic mining offers an unparalleled view of microbial diversity, enabling the discovery of novel genes from the vast uncultured majority. For research focused on the stability and function of extremophile enzymes, a synergistic approach is most powerful. Metagenomics can guide the targeted cultivation of elusive organisms and rapidly identify promising genetic targets, while traditional culturing remains essential for the experimental characterization that confirms an enzyme's stability and industrial potential.
The term "microbial dark matter" (MDM) describes the enormous fraction of microorganisms that refuse to grow in standard laboratory cultures, representing an estimated >95% of prokaryotic lineages in environments like marine sediments and hydrothermal vents [37]. This vast, unexplored biological terrain constitutes a hidden reservoir of novel enzymes, metabolic pathways, and biochemical functions with immense potential for biotechnology and drug development [14] [38]. Overcoming the challenge of MDM requires advanced, culture-independent techniques that can link genetic potential to biological function, enabling researchers to tap into this untapped resource for discovering robust industrial enzymes, including those from extremophiles [37] [39].
Two primary methodological paradigms have emerged to illuminate this microbial dark matter: single-cell genomics (SCG), which isolates and sequences the DNA of individual cells, and function-based screening, which directly assays metagenomic libraries for desired enzymatic activities [40] [41]. This guide provides a comparative analysis of these core approaches, detailing their respective workflows, capabilities, and limitations. It is framed within a broader thesis on the comparative stability of enzymes, with a specific focus on the unique value of extremophile-derived enzymes, known for their extraordinary stability under industrial harsh conditions [14] [39].
Comparative Workflows: Two Pathways to Discovery
The following diagram illustrates the core workflows for single-cell genomics and function-based metagenomic screening, highlighting their distinct pathways from environmental sample to functional discovery.
Single-Cell Genomics: Illuminating Genomic Blueprints
Core Methodology and Experimental Protocol
Single-cell genomics enables researchers to access the genetic material of individual uncultured cells directly from environmental samples. The standard workflow, as derived from the search results, involves several critical steps [40] [42]:
- Sample Preparation and Preservation: Environmental samples (e.g., biofilm, sludge, sediment) are collected and preserved to maintain cellular integrity, often using fixatives like ethanol or glutaraldehyde.
- Single-Cell Isolation: This is a crucial step where individual cells are separated from the complex community. While Fluorescence-Activated Cell Sorting (FACS) is most common, a novel label-free single-cell dispensing system offers advantages. This microfluidic printer captures cells in picoliter droplets, assesses cell number and morphology via automated light microscopy, and deposits single cells into wells with minimal stress [42].
- Cell Lysis and Whole Genome Amplification: The membrane of the isolated cell is chemically or enzymatically lysed. The minimal genomic DNA is then amplified using methods like Multiple Displacement Amplification (MDA) to generate sufficient material for sequencing [40].
- Screening and Sequencing: Amplified genomes are often screened using 16S rRNA gene PCR to phylogenetically identify the captured cell. Subsequently, library preparation (e.g., with Illumina Nextera XT) and sequencing are performed to generate a Single Amplified Genome (SAG) [40] [42].
Performance and Key Experimental Data
Recent studies demonstrate the power of SCG in capturing novel microbial diversity. The table below summarizes quantitative data from a key study that utilized the novel single-cell dispensing approach [42].
Table 1: Genome Recovery Success Rates for Single-Cell Genomics
| Metric | Performance Data | Experimental Context |
|---|---|---|
| Genome Recovery Rate | 81.5% (717 out of 880 sorted cells) | Label-free single-cell dispensing from wastewater [42] |
| 16S rRNA Analysis Success | 50.1% of amplified cells (359 out of 717) | Successful phylogenetic identification post-amplification [42] |
| Novel SAGs Recovered | 27 Single Amplified Genomes | Representing 15 novel Patescibacteria/CPR species, genera, and families [42] |
| Complementarity to Metagenomics | High (Recovery of lineages missed by MAGs) | Phylogenetically distinct SAGs were not captured by genome-resolved metagenomics of the same sample [42] |
Function-Based Screening: Direct Discovery of Novel Enzymes
Core Methodology and Experimental Protocol
Function-based metagenomic screening bypasses sequencing and directly tests for desired enzymatic activities, enabling the discovery of entirely novel genes with no similarity to known sequences [41]. The general workflow is as follows [41] [37]:
- Metagenomic Library Construction: Total environmental DNA is extracted and fragmented. These fragments are cloned into suitable expression vectors (plasmids, fosmids, or cosmids) and introduced into a surrogate host, typically Escherichia coli.
- Activity-Based Screening: The library of clones is subjected to an assay designed to detect the target activity. This is the core of the functional approach.
- Hit Identification and Validation: Clones exhibiting a positive signal in the assay are selected. The metagenomic insert is sequenced to identify the gene responsible for the activity, which is then characterized biochemically.
The screening step employs various strategies, each with specific protocols and detection methods:
- Agar Plate Screening: The simplest and most common method. Clones are grown on agar plates containing a substrate that produces a visual change upon enzymatic activity.
- Microtiter Plate Screening: A higher-throughput method where clones are grown in microtiter plates, and activity is measured spectrophotometrically or fluorometrically (e.g., using p-nitrophenyl derivatives) [41].
- Ultra-High-Throughput Screening (uHTS): Advanced methods like FACS-based screening and droplet-based microfluidics are used to screen libraries of immense size (e.g., >10^9 clones). These systems encapsulate single cells in water-in-oil droplets together with a fluorescent substrate, enabling the sorting of millions of cells per hour based on activity [41].
Performance and Key Experimental Data
Function-based screening has successfully identified a wide range of novel enzymes from diverse environments. The data below highlights its application and effectiveness.
Table 2: Representative Enzymes Discovered via Function-Based Screening
| Target Enzyme | Detection Method/Substrate | Source Environment | Key Finding |
|---|---|---|---|
| Protease | Phenotypical detection / 1% skim-milk | Goat skin, desert sands, forest soil | Discovery of novel proteases active against insoluble protein substrates [41] |
| Esterase/Lipase | Phenotypical detection / 1% Tributyrin | Marine mud, sponge, alluvial soil | Identification of esterases and lipases with activity in high-salt or low-temperature conditions [41] |
| γ-Lactamase | Ninhydrin stain on filter overlay | Sulfolobus solfataricus (thermophile) | Discovery of a thermostable enzyme critical for synthesizing the anti-HIV drug Abacavir [39] |
| L-Aminoacylase | -- | Thermococcus litoralis (thermophile) | Identification of a thermostable enzyme with broad substrate specificity for unnatural amino acids [39] |
Comparative Analysis: Strengths, Limitations, and Synergy
The following table provides a direct comparison of the two methodologies, contextualizing their performance and utility for researchers.
Table 3: Direct Comparison of Single-Cell Genomics and Function-Based Screening
| Aspect | Single-Cell Genomics (SCG) | Function-Based Screening |
|---|---|---|
| Primary Output | Single Amplified Genomes (SAGs) - Genomic context of individual cells [42] | Novel enzyme genes with confirmed activity [41] |
| Key Strength | Accesses rare biosphere and cell-cell associations; provides full genomic context [42] | Discovers completely novel enzymes without prior sequence knowledge (functional dark matter) [41] [43] |
| Major Limitation | Genome amplification can be incomplete/biased; high cost [44] | Low hit rates; dependent on successful expression in a heterologous host [41] [37] |
| Throughput | Medium (hundreds to thousands of cells per run) [42] | Very High (millions of clones with uHTS) [41] |
| Ideal for Research on | Microbial ecology, evolution, and uncovering novel phyla (e.g., Patescibacteria) [42] [44] | Industrial enzyme discovery, particularly for biocatalysis and therapeutics [41] [39] |
| Role in Stability Research | Identifies genetic adaptations (e.g., in extremophiles) that confer stability at the genomic level [14] | Directly tests the stability and function of discovered enzymes under process-like conditions (e.g., high T, pH) [39] |
The Scientist's Toolkit: Essential Research Reagents and Solutions
Successful implementation of these advanced methodologies relies on a suite of specialized reagents and tools. The table below lists key solutions for researchers embarking on MDM exploration.
Table 4: Essential Research Reagent Solutions for Microbial Dark Matter Studies
| Reagent / Solution | Function / Application | Relevant Context |
|---|---|---|
| Fluorescence-Activated Cell Sorter (FACS) | High-throughput, staining-based sorting of single cells from environmental samples for SCG [40] | Standard method for single-cell isolation; can cause cell stress [42] |
| Microfluidic Single-Cell Dispenser | Label-free, image-based isolation of single cells, minimizing stress for downstream genomics or cultivation [42] | An alternative to FACS; allows morphological selection [42] |
| Multiple Displacement Amplification (MDA) Kit | Isothermal whole-genome amplification of minute DNA amounts from a single cell [40] | Critical for SCG to generate sufficient DNA for sequencing from one cell [40] [42] |
| Fosmids / Cosmids | Large-insert cloning vectors for constructing metagenomic libraries, capturing large gene clusters [41] [37] | Essential for function-based screening to maintain large DNA fragments and operons [41] |
| Activity-Specific Substrates (e.g., AZCL-casein, Tributyrin) | Chromogenic or fluorogenic substrates used in agar or microtiter plate assays to detect enzymatic activity [41] | Enable visual detection of enzyme hits in functional metagenomic screens [41] |
| Nextera XT DNA Library Prep Kit | Preparation of sequencing-ready libraries from amplified genomic DNA for Illumina platforms [40] | Standardized protocol for sequencing SAGs and other low-input samples [40] |
Single-cell genomics and function-based metagenomic screening are powerful, complementary technologies in the campaign to overcome the challenge of microbial dark matter. SCG excels at revealing the genomic blueprint and ecological context of uncultured organisms, including extremophiles whose genetic adaptations hint at the stability of their encoded enzymes [14] [42]. In contrast, function-based screening directly uncovers novel enzymatic activities, even from genes with no known homologs, providing a direct path to robust biocatalysts like extremozymes that are already functional under industrial process conditions [41] [39].
For researchers focused on the comparative stability of microbial versus extremophile enzymes, a synergistic approach is most powerful. SCG can identify extremophile lineages and their unique genetic signatures, while functional screening can validate the stability and catalytic prowess of the enzymes these genes encode. By integrating these methodologies, the scientific community can systematically illuminate microbial dark matter, transforming it from a biological enigma into a boundless resource for drug development and industrial biotechnology.
The increasing demand for industrial enzymes that can withstand harsh process conditions—such as extreme temperatures, pH, and organic solvents—has driven significant interest in extremozymes derived from microorganisms inhabiting Earth's most challenging environments [3] [45]. These enzymes, native to extremophiles, offer superior activity and stability under non-standard conditions compared to their mesophilic counterparts, making them highly desirable for applications in biotechnology, pharmaceuticals, and bioremediation [46]. However, a major challenge impedes their widespread adoption: the difficulty of cultivating the source extremophilic organisms on an industrial scale due to their fastidious growth requirements and low biomass yields [45].
Heterologous expression in mesophilic hosts presents a powerful solution to this bottleneck. This strategy involves cloning the extremozyme-encoding gene from the extremophile into a robust, easily cultivated mesophilic host such as Escherichia coli or Bacillus subtilis, enabling large-scale production of the enzyme under standard laboratory and industrial conditions [46]. This guide objectively compares the leading mesophilic host systems for extremozyme production, providing experimental data and methodologies to inform researchers and drug development professionals in their selection of an appropriate expression platform.
Comparing Mesophilic Host Systems for Extremozyme Production
The success of heterologous expression hinges on selecting an appropriate host-vector system. The table below compares the key characteristics of the most commonly employed mesophilic hosts for extremozyme production.
Table 1: Comparison of Mesophilic Host Systems for Heterologous Extremozyme Production
| Host System | Optimal Expression Vector Features | Key Advantages | Major Limitations | Ideal for Extremozyme Types |
|---|---|---|---|---|
| Escherichia coli | Vectors with T5/lac promoter [46], pUC origin (high copy) [47], RSF origin (stable medium copy) [47]. | Rapid growth, high transformation efficiency, well-characterized genetics, high protein yields [46]. | Improper folding of complex proteins, formation of inclusion bodies, lack of post-translational modifications [48]. | Thermophilic enzymes [48], psychrophilic catalases [46], amine-transaminases [46]. |
| Bacillus subtilis | Shuttle vectors (e.g., pUB110), integration vectors for stability, optimized promoters/RBS [49]. | Efficient protein secretion, GRAS status, no endotoxin production [49]. | Poor plasmid stability, low copy number, protease activity degrading target protein [49]. | Secreted extremozymes (e.g., proteases, amylases), industrial-scale enzyme production [49]. |
| Other Engineered Hosts | Species-specific replicons and promoters (e.g., for Thermus thermophilus) [48]. | Can be grown at elevated temperatures, aiding correct folding of thermophilic enzymes [48]. | Limited genetic tools, often requires gene synthesis due to high GC content, slower growth [48]. | Complex thermophilic archaeal enzymes that misfold in E. coli [48]. |
Experimental Workflow for Extremozyme Expression and Characterization
Developing a recombinant extremozyme product involves a multi-phase process from discovery to scaled-up production. The following workflow and detailed protocols are adapted from established methods for producing catalases, laccases, and amine-transaminases from extremophiles [46].
Diagram 1: A generalized experimental workflow for the heterologous production of extremozymes, from discovery in environmental samples to scaled-up production.
Phase 1: Discovery of Extremophiles and Target Gene Isolation
Protocol 1: Functional Screening for Thermophilic Laccase Producers
- Sample Inoculation: Inoculate environmental samples (e.g., from geothermal hot springs) into liquid culture media. Incubate at 50°C, pH 8.0, with shaking for 24-48 hours. Supplement media with 0.1% lignin to induce laccase gene expression [46].
- Activity Screening: Plate serial dilutions of the enrichment culture on solid agar containing 0.5 mM guaiacol. Laccase-positive colonies will appear surrounded by a brown halo due to the oxidation of guaiacol [46].
- Strain Identification: Purify positive colonies through repeated streaking. Identify the isolate using 16S rRNA gene sequencing and whole-genome sequencing to locate the laccase-encoding gene [46].
Protocol 2: Gene Cloning Strategy
- PCR Amplification: Design primers specific to the identified laccase gene. Amplify the coding sequence using high-fidelity DNA polymerase from the extremophile's genomic DNA [46].
- Vector Ligation: Clone the purified PCR product into an expression vector (e.g., one featuring a T5 promoter and kanamycin resistance). Use ligation-independent cloning or restriction enzyme-based methods [46].
- Transformation: Introduce the constructed plasmid into a competent mesophilic host, typically E. coli BL21(DE3) strains, via heat-shock or electroporation. Select transformants on agar plates containing the appropriate antibiotic [46].
Phase 2: Heterologous Expression and Purification
Protocol 3: Small-Scale Expression and Cell Lysis
- Starter Culture: Inoculate a single transformed colony into 5-10 mL of LB medium with antibiotic. Grow overnight at 37°C with shaking at 180 rpm [46].
- Expression Culture: Dilute the overnight culture 1:100 into a fresh, larger volume of medium. Grow at 37°C until the optical density at 600 nm (OD₆₀₀) reaches 0.6-0.8 [46].
- Protein Induction: Add Isopropyl β-d-1-thiogalactopyranoside to a final concentration of 0.1-0.5 mM to induce gene expression. For challenging proteins, reduce the temperature to 25-30°C and extend the induction time to 6-16 hours to improve soluble yield [46].
- Cell Harvest: Centrifuge the culture at >9,000 × g for 15 minutes at 4°C. Discard the supernatant and resuspend the cell pellet in an appropriate lysis buffer (e.g., 50 mM Tris-HCl, pH 8.0, 150 mM NaCl) [46].
- Cell Disruption: Lyse the cells using sonication (e.g., ten cycles of 15-second pulses with 45-second rests on ice) or high-pressure homogenization. Remove cell debris by centrifugation at >14,000 × g for 30 minutes at 4°C. The resulting supernatant is the soluble crude extract [46].
Phase 3: Biochemical Characterization and Stability Assessment
A critical step is comparing the performance of the recombinant extremozyme to commercial mesophilic enzymes. The following table and protocol outline a standard characterization.
Table 2: Comparative Biochemical Characterization of a Recombinant Thermophilic Laccase vs. a Commercial Mesophilic Laccase
| Enzyme Parameter | Experimental Condition | Recombinant Thermophilic Laccase | Commercial Mesophilic Laccase |
|---|---|---|---|
| Optimal Temperature | Temperature gradient (20-90°C) | 70°C [50] | 40°C |
| Thermal Stability | Incubation at 60°C for 1 hour | >90% residual activity [50] | <10% residual activity |
| Optimal pH | pH gradient (3.0-9.0) | pH 8.0 [46] | pH 6.0 |
| Half-life at 70°C | Continuous incubation at 70°C | >4 hours [50] | Not stable |
| Activity in Solvents | 10% (v/v) Methanol | 85% residual activity | 45% residual activity |
Protocol 4: Temperature Optimum and Thermostability Assay
- Temperature Optimum: Perform the standard enzyme activity assay at temperatures ranging from 20°C to 90°C. The activity can be measured spectrophotometrically by monitoring the oxidation of a substrate like 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) at 420 nm [50]. Plot the relative activity against temperature to determine the optimum.
- Thermostability: Incubate the enzyme solution at a high temperature (e.g., 60°C or 70°C). Withdraw aliquots at regular time intervals (e.g., 0, 15, 30, 60, 120 minutes), cool them immediately on ice, and measure the residual activity under standard assay conditions. Calculate the half-life from the decay of activity over time [50].
The Scientist's Toolkit: Essential Research Reagents
The table below lists key reagents and materials required for establishing a heterologous expression pipeline for extremozymes.
Table 3: Essential Research Reagents for Heterologous Extremozyme Production
| Reagent/Material | Function/Purpose | Specific Examples & Notes |
|---|---|---|
| Expression Vectors | Carries the gene of interest and enables its controlled expression in the host. | Vectors with T5/lac promoters [46]; pUC (high-copy) [47] or RSF (stable medium-copy) origins [47]. |
| Competent Cells | Mesophilic host cells rendered permeable for DNA uptake. | E. coli BL21(DE3) for protein expression; B. subtilis strains like 168 for secretion [49]. |
| Inducers | Triggers transcription of the target gene. | Isopropyl β-d-1-thiogalactopyranoside for lac-based systems [46]. |
| Culture Media | Supports the growth of the mesophilic host. | Luria-Bertani or Terrific Broth for E. coli; specialized media for other hosts. |
| Lysis Reagents | Breaks open host cells to release the recombinant protein. | BugBuster reagent [47] or lysozyme; buffers for sonication/homogenization. |
| Chromogenic Substrates | Allows for visual or spectroscopic detection of enzyme activity. | Guaiacol (for laccases) [46]; ABTS [50]; substrate analogs for other enzyme classes. |
The heterologous expression of extremozymes in mesophilic hosts is a cornerstone strategy for bridging the gap between the discovery of novel biocatalysts and their industrial application. As demonstrated, *E. coli remains the dominant workhorse* due to its unparalleled ease of use and yield for many proteins [46]. However, for enzymes requiring complex folding or secretion, *B. subtilis and specialized thermophilic hosts* present viable alternatives [49] [48].
Future advancements will likely be driven by synthetic biology, with the creation of more sophisticated expression vectors and engineered mesophilic chassis capable of performing host-specific post-translational modifications [48] [49]. Furthermore, the integration of machine learning for predicting expression and solubility based on gene sequence will streamline the production of challenging extremozymes [50]. By leveraging and continually improving these heterologous systems, researchers can fully harness the power of extremophiles to develop robust biocatalysts that meet the demanding needs of modern biotechnology and drug development.
L-Asparaginase (L-ASNase) is a crucial therapeutic enzyme used in the treatment of acute lymphoblastic leukemia (ALL) and other hematopoietic malignancies. The enzyme exerts its antineoplastic effect by hydrolyzing circulating L-asparagine into aspartic acid and ammonia, depleting this essential amino acid required by malignant lymphoblasts that lack sufficient expression of asparagine synthetase (ASNS). Current commercial formulations, primarily derived from Escherichia coli and Erwinia chrysanthemi, face significant limitations including hypersensitivity reactions, development of neutralizing antibodies, short half-life, and undesirable glutaminase co-activity that contributes to neurotoxicity [51].
The search for novel L-ASNase variants with improved therapeutic profiles has led researchers to explore extremophilic microorganisms as promising enzyme sources. Microbes thriving in extreme environments have evolved unique enzymatic adaptations, resulting in proteins with enhanced stability, novel structural features, and distinct immunological properties [14]. This case study provides a comprehensive comparative analysis of a novel Type II L-ASNase from halotolerant Bacillus subtilis CH11 isolated from Chilca salterns in Peru, evaluating its potential as a therapeutic alternative against current commercial formulations [52].
The Candidate: Type II L-ASNase from HalotolerantBacillus subtilisCH11
Source and Production
The Bacillus subtilis CH11 strain was isolated from the hypersaline Chilca salterns in Lima, Peru, representing a halotolerant organism adapted to saline conditions. The genome of B. subtilis contains two L-ASNase encoding genes: ansA (type I, cytosolic, low substrate affinity) and ansZ (type II, extracellular, higher substrate affinity) [52]. Researchers cloned an N-terminally truncated version of the ansZ gene (designated ansZP21, lacking the first 60 base pairs encoding the signal peptide YccC) into a pET-15b expression vector and expressed it heterologously in E. coli BL21(DE3)pLysS [52] [53].
The recombinant enzyme (L-ASNaseZP21) was purified using immobilized metal affinity chromatography (IMAC) with a one-step nickel affinity system, yielding a protein with specific activity of 234.38 U/mg. Size exclusion chromatography revealed the native enzyme exists as a homotetramer with an estimated molecular weight of 155 kDa [52].
Bioinformatic Characterization
Bioinformatic analysis using SignalP-6.0 confirmed the removal of the signal peptide, while ProtParam was employed to predict molecular weight and isoelectric point. Structural predictions were performed using AlphaFold2, providing insights into the monomeric structure [52] [53].
Comparative Performance Analysis
Kinetic Parameters
Kinetic parameters are critical determinants of therapeutic efficacy, with low Km (high substrate affinity) being particularly important for efficient asparagine depletion at physiological concentrations (approximately 50 μM) [51].
Table 1: Comparative Kinetic Parameters of Therapeutic L-Asparaginases
| Enzyme Source | Km (μM) | Vmax (μmol/mL/min) | Kcat (s⁻¹) | Specific Activity (U/mg) |
|---|---|---|---|---|
| Bacillus subtilis CH11 (L-ASNaseZP21) | 4,752 μM | 145.2 | - | 234.38 [52] |
| Chryseomicrobium amylolyticum (CamASNase) | 6.364 | 909.09 | 222.82 | 173.69 [54] |
| Commercial Benchmarks | ||||
| E. coli (Oncaspar) | ~50 | - | 200-560 | - [51] |
| Erwinia chrysanthemi (Erwinase) | ~50 | - | 200-560 | - [51] |
The B. subtilis CH11 enzyme demonstrates a substantially higher Km value (4.75 mM) compared to both commercial formulations (∼50 μM) and the recently characterized C. amylolyticum enzyme (6.364 μM) [52] [51] [54]. This indicates lower substrate affinity, which may impact its therapeutic efficiency at physiological asparagine concentrations.
Biochemical and Stability Properties
Biochemical characterization under various conditions provides insights into practical utility and formulation requirements.
Table 2: Comparative Biochemical Properties of L-Asparaginases
| Property | B. subtilis CH11 | C. amylolyticum | Commercial Formulations |
|---|---|---|---|
| Optimal pH | 9.0 [52] | - | - |
| Optimal Temperature | 60°C [52] | - | - |
| Thermal Stability | Half-life: 3h 48min at 60°C; retains 50% activity for 24h at 37°C [52] | Serum half-life: ~27 minutes [54] | Varies; PEGylation extends half-life [51] |
| Effect of Modulators | Enhanced by KCl, CaCl2, MgCl2, mercaptoethanol, DL-dithiothreitol [52] | - | - |
| Glutaminase Activity | Not specified | Absent [54] | Present in E. coli and Erwinia enzymes (∼5%) [51] |
| Urease Activity | Not specified | Absent [54] | Varies |
The B. subtilis CH11 enzyme exhibits remarkable thermostability with optimal activity at 60°C and retains 50% of its initial activity for 24 hours at physiological temperature (37°C), indicating good stability under treatment conditions [52]. Its activity enhancement by various ions and reducing agents suggests potential compatibility with physiological milieus.
Structural Comparison
Structural features significantly influence enzyme stability, immunogenicity, and functional efficiency.
Table 3: Structural Characteristics of L-Asparaginases
| Structural Feature | B. subtilis CH11 | C. amylolyticum | Commercial Formulations |
|---|---|---|---|
| Quaternary Structure | Homotetramer (155 kDa) [52] | - | Varies; often tetrameric |
| Sequence Identity | - | Low identity with commercial enzymes [54] | Reference standards |
| Allergenic Potential | - | Predicted nonallergenic and nontoxic [54] | Immunogenic in some patients [51] |
The homotetrameric structure of the B. subtilis CH11 enzyme is consistent with many bacterial L-ASNases. The low sequence identity of novel enzymes like C. amylolyticum with commercial formulations suggests reduced potential for cross-reactive immunogenicity, a significant advantage for patients who develop hypersensitivity to current therapeutics [51] [54].
Experimental Protocols for Key Analyses
Enzyme Expression and Purification Protocol
Cloning and Expression [52] [53]:
- Amplify the ansZP21 gene (N-terminally truncated ansZ) from B. subtilis CH11 genomic DNA using specific primers with NdeI and BamHI restriction sites
- Digest PCR product and pET-15b vector with restriction enzymes
- Ligate insert into vector using T4 DNA ligase
- Transform into E. coli DH5α and verify correct cloning by sequencing
- Transform expression vector into E. coli BL21(DE3)pLysS
- Grow culture in LB-Miller medium with 100 μg/mL ampicillin at 37°C until OD600 reaches 0.6
- Induce expression with 0.5 mM IPTG and incubate for 14 hours at 22°C
Purification [52]:
- Harvest cells by centrifugation (2,133 × g, 20 minutes, 4°C)
- Disrupt cells using BugBuster Master Mix reagent
- Recover clarified lysate by centrifugation (12,555 × g, 30 minutes, 4°C)
- Purify using FPLC with HisTrap FF column pre-equilibrated with 50 mM Tris-HCl, 100 mM NaCl, 20 mM imidazole, pH 8.5
- Wash with 5 column volumes of equilibration buffer
- Elute with linear imidazole gradient (up to 500 mM)
- Desalt in Tris-HCl pH 8.5 and store at 4°C
Enzyme Activity and Kinetic Characterization
Activity Assay (Nessler's Method) [52] [53]:
- Prepare reaction mixture: 100 μL of 50 mM Tris-HCl pH 8.6, 10 μL of 189 mM L-asparagine, 90 μL H2O, and 10 μL enzyme sample
- Incubate for 10 minutes at 37°C
- Stop reaction by adding 10 μL trichloroacetic acid
- Add 140 μL Nessler's reagent and measure absorbance at 505 nm
- Calculate activity using ammonium sulfate standard curve
- One unit of enzyme activity defined as amount producing 1 μmol ammonia per minute under assay conditions
Kinetic Parameter Determination [52]:
- Measure initial reaction rates at varying L-asparagine concentrations (0.1-10 mM)
- Plot reaction velocity versus substrate concentration
- Determine Km and Vmax using Michaelis-Menten equation nonlinear regression
Zymography [53]:
- Separate proteins by native PAGE
- Incubate gel in solution containing 50 mM Tris-HCl pH 8.6, 189 mM L-asparagine, 2 M hydroxylamine, 2 M NaOH at 37°C for 20 minutes
- Stain with solution containing 10% FeCl3, 5% trichloroacetic acid, 0.66 M HCl
- Visualize L-ASNase activity bands based on L-aspartic acid β-hydroxamate formation
Research Reagent Solutions
Table 4: Essential Research Reagents for L-Asparaginase Characterization
| Reagent/Chemical | Function/Application | Specific Example |
|---|---|---|
| pET-15b Vector | Expression vector with N-terminal His-tag | Novagen [52] |
| E. coli BL21(DE3)pLysS | Expression host for recombinant protein | Thermo Scientific [52] |
| BugBuster Master Mix | Cell lysis and protein extraction | Novagen [52] |
| HisTrap FF Column | Immobilized metal affinity chromatography | GE Healthcare [52] |
| Nessler's Reagent | Ammonia detection for activity assay | Sigma-Aldrich [52] [55] |
| IPTG | Induction of protein expression | Sigma-Aldrich [52] |
| Phusion DNA Polymerase | High-fidelity PCR amplification | Thermo Scientific [52] |
| Restriction Enzymes (NdeI, BamHI) | Vector and insert digestion | New England Biolabs [52] |
Therapeutic Potential and Research Implications
Cytotoxicity and Selectivity
While the B. subtilis CH11 enzyme requires further cytotoxicity assessment, the C. amylolyticum L-ASNase demonstrates promising antineoplastic activity with IC50 values of 0.043, 0.041, and 0.05 IU/mL against MCF7, MOLT4, and THP-1 cell lines, respectively. Importantly, it showed no toxicity toward normal human fibroblast cells and was non-hemolytic, indicating favorable selectivity [54]. Flow cytometry analysis revealed 81.4% apoptotic cells in MOLT-4 cells at IC50, confirming potent anticancer activity through apoptosis induction [54].
Advantages and Limitations
The B. subtilis CH11 L-ASNase offers several advantageous features including high thermostability, compatibility with physiological additives, and robust production yield. However, its relatively high Km (4.75 mM) compared to commercial formulations (~50 μM) and emerging alternatives (6.364 μM for C. amylolyticum) represents a significant limitation for therapeutic application, where high affinity is essential for efficient asparagine depletion at physiological concentrations (~50 μM) [52] [51] [54].
Future Research Directions
Future studies should focus on:
- Protein engineering to enhance substrate affinity while maintaining stability
- Comprehensive in vivo efficacy and toxicity assessments
- Immunogenicity profiling compared to current formulations
- Half-life extension strategies, including PEGylation or nanoparticle immobilization
- Scaling up production using optimized bioreactor conditions
Visualizations
Comparative Analysis Workflow
Experimental Process Flow
This comparative analysis demonstrates that while the Type II L-asparaginase from halotolerant Bacillus subtilis CH11 exhibits promising biochemical characteristics, particularly regarding thermostability and production yield, its high Km presents a significant challenge for direct therapeutic application. The enzyme's performance highlights both the potential and limitations of extremophile-derived enzymes for cancer therapy. Future research should focus on protein engineering approaches to enhance substrate affinity while maintaining the advantageous stability properties of this enzyme. The growing pipeline of novel L-ASNases from diverse microbial sources, including those from extreme environments, continues to expand the possibilities for developing improved therapeutics with reduced side effects and enhanced efficacy for cancer patients.
Industrial biocatalysis leverages enzymes as selective and efficient catalysts for manufacturing complex molecules, particularly in the pharmaceutical industry. Among these, γ-lactamases and robust proteases represent two critical classes of biocatalysts enabling the synthesis of vital therapeutics and functioning under industrial process conditions. γ-Lactamases specifically catalyze the enantioselective hydrolysis of racemic γ-lactam (2-azabicyclo[2.2.1]hept-5-en-3-one), a key step in producing optically pure (−)-γ-lactam, which serves as the essential building block for antiviral agents such as abacavir and carbovir [56] [57]. Concurrently, proteases derived from extremophilic microorganisms exhibit remarkable stability under harsh conditions, making them invaluable for applications requiring prolonged catalyst lifetime and operational robustness [14] [6]. This guide provides a comparative analysis of the performance and stability of microbial and extremophile-derived enzymes, focusing on γ-lactamases for antiviral drug synthesis and robust proteases for industrial applications, supported by experimental data and methodologies.
γ-Lactamases for the Synthesis of Antiviral Drugs
Function and Industrial Significance
γ-Lactamases enable the kinetic resolution of racemic γ-lactam by selectively hydrolyzing one enantiomer, leaving behind the desired optically pure counterpart. The (−)-γ-lactam is a critical synthon for carbocyclic nucleoside analogues, a class of compounds that includes potent anti-HIV and antiviral medications [56] [57]. The enzymatic route provides a more efficient and environmentally friendly alternative to chemical synthesis for obtaining these high-value chiral intermediates [56].
Comparative Performance of Microbial γ-Lactamases
Enzymes from diverse microbial sources have been characterized, showing variations in activity, enantioselectivity, and stability. The table below summarizes key biochemical properties of several documented γ-lactamases.
Table 1: Comparative Biochemical Properties of Characterized γ-Lactamases
| Enzyme Source | Enantiopreference | Optimal Temperature (°C) | Optimal pH | Key Catalytic Features/Remarks | Reference |
|---|---|---|---|---|---|
| Microbacterium hydrocarbonoxydans (Mh33H4-5540) | (+)-γ-lactam | 25 | 8.0 | Activity boosted 5.5-fold by Ni²⁺ or Co²⁺; novel isochorismatase-like hydrolase family. | [58] |
| Aureobacterium species | (−)-γ-lactam | Information Missing | Information Missing | Belongs to α/β-hydrolase fold family; features a classic Ser-His-Asp catalytic triad. | [57] |
| Comamonas acidovorans | (+)-γ-lactam | Information Missing | Information Missing | High sequence identity to formamidases/acetamidases; structure solved (PDB: 2WKN). | [56] |
| Aeropyrum pernix | (+)-γ-lactam | Information Missing | Information Missing | Thermostable type II (+)-γ-lactamase from hyperthermophilic archaeon. | [58] |
Experimental Protocol for γ-Lactamase Activity Assay
The following methodology is commonly employed to determine γ-lactamase activity and enantioselectivity, crucial for evaluating their suitability for industrial resolution processes [56] [58].
- Enzyme Purification: Heterologously express the target γ-lactamase gene (e.g., in E. coli). Purify the enzyme to homogeneity using chromatography techniques such as nickel-affinity chromatography for His-tagged proteins.
- Hydrolysis Reaction Setup: Incubate the purified enzyme with a racemic mixture of γ-lactam (e.g., 10 mM) in a suitable buffer (e.g., Tris-HCl, pH 8.0) at the enzyme's optimal temperature (e.g., 25°C).
- Reaction Monitoring: Terminate the reaction at different time points by heat inactivation or acidification.
- Product Analysis:
- Chromatographic Separation: Analyze the reaction mixture using chiral High-Performance Liquid Chromatography (HPLC) or Gas Chromatography (GC) to separate and quantify the remaining (−)-γ-lactam and the formed (+)-γ-amino acid.
- Enantiomeric Excess (ee) Calculation: Determine the enantiomeric excess using the formula: ( ee (\%) = \frac{([S] - [R])}{([S] + [R])} \times 100 ), where [S] and [R] are the concentrations of the two enantiomers.
- Yield Calculation: Calculate the chemical yield of the desired enantiomer.
- Kinetic Parameter Determination: Measure initial reaction rates at varying substrate concentrations. Plot these rates to calculate Michaelis-Menten constants ((Km)) and maximum velocity ((V{max})) using Lineweaver-Burk or non-linear regression analysis.
Diagram 1: Workflow for determining γ-lactamase activity and enantioselectivity.
Robust Proteases: Microbial vs. Extremophile Enzymes
Stability as a Key Performance Indicator
For industrial biocatalysis, enzyme stability under operational conditions (e.g., high temperature, extreme pH, organic solvents) is as critical as activity. Extremophile-derived enzymes, or extremozymes, naturally exhibit superior stability due to structural adaptations, making them promising candidates for harsh industrial processes [14] [6].
Comparative Analysis of Protease Stability
The table below compares the stability features and industrial advantages of proteases from extremophiles versus general microbial sources.
Table 2: Stability Comparison of Microbial and Extremophile-Derived Proteases
| Enzyme Source Type | Thermal Stability | pH Stability | Structural Adaptations | Industrial Advantages |
|---|---|---|---|---|
| Psychrophiles (Cold-adapted) | Low optimal temperature; inactivated by moderate heat. | Varies, often narrow. | Increased molecular flexibility, reduced proline/arginine, surface-loaded residues [14]. | Energy-saving, low-temperature processing. |
| Thermophiles (e.g., Thermus sp.) | High optimal temperature (up to 100°C); prolonged half-life at 60-80°C [14]. | Varies, often broad. | Compact oligomers, reinforced hydrophobic cores, disulfide bonds, ionic networks [14]. | Resistance to contamination, high substrate solubility, reduced viscosity. |
| Halophiles (Salt-loving) | Moderate to high, often linked to salt concentration. | Varies. | Acidic surface residues, solvation shell maintenance in low-water activity [6]. | Functionality in high-salt or low-water environments. |
| General Microbial (Mesophilic) | Moderate; optimal 20-45°C. | Varies, can be narrow. | Standard folded structures. | Well-established production; may require stabilization. |
Enzyme Stabilization Strategies for Industrial Applications
Enzyme immobilization is a primary technology to enhance stability, reusability, and to facilitate easy separation from reaction mixtures, thereby reducing operational costs [11]. The choice of method depends on the enzyme and the application requirements.
Table 3: Common Enzyme Immobilization Techniques and Their Characteristics
| Immobilization Method | Mechanism of Binding | Advantages | Disadvantages |
|---|---|---|---|
| Adsorption | Weak forces (van der Waals, ionic, hydrophobic) [11]. | Simple, reversible, high activity retention, low cost [11]. | Enzyme leakage under changing pH/ionic strength [11]. |
| Covalent Binding | Covalent bonds between enzyme and activated support [11]. | No enzyme leakage, high stability, reusable support [11]. | Potential activity loss, complex process, higher cost [11]. |
| Encapsulation / Entrapment | Physical confinement within a porous matrix [11]. | Protection from denaturants and proteases. | Mass transfer limitations, enzyme leaching. |
| Bacterial Spore Surface Display | Genetic fusion or adsorption to spore coat proteins [59]. | GRAS status, high natural stability, integrated production/immobilization [59]. | Relatively low display efficiency, requires genetic engineering [59]. |
Experimental Protocol for Enzyme Immobilization and Stability Assessment
The following protocol outlines steps for covalent immobilization and subsequent stability testing, a widely used method for creating robust biocatalysts [11].
- Support Activation:
- Incubate the chosen support material (e.g., chitosan, porous silica) with a cross-linking agent (e.g., glutaraldehyde) in buffer.
- Wash the activated support thoroughly to remove excess reagent.
- Enzyme Immobilization:
- Mix the purified enzyme solution with the activated support.
- Incubate for several hours under mild shaking to allow covalent coupling.
- Recover the immobilized enzyme by filtration or centrifugation and wash extensively to remove unbound enzyme.
- Activity Recovery Measurement:
- Assay the activity of the immobilized enzyme and compare it to an equivalent amount of free enzyme before immobilization.
- Calculate the activity recovery percentage.
- Thermal Stability Assay:
- Incubate both free and immobilized enzymes at a elevated temperature (e.g., 60°C).
- Withdraw samples at regular time intervals and measure residual activity.
- Plot residual activity vs. time to determine the half-life of each form.
- Reusability Test:
- Use the immobilized enzyme for multiple consecutive reaction cycles.
- After each cycle, recover the immobilized enzyme, wash it, and assay its activity for the next cycle.
- Plot the residual activity versus the number of cycles.
Diagram 2: Experimental workflow for enzyme immobilization and stability assessment.
The Scientist's Toolkit: Key Research Reagent Solutions
This table lists essential materials and reagents used in the experiments cited within this guide, along with their primary functions.
Table 4: Essential Reagents for Enzyme Biocatalysis Research
| Reagent / Material | Function / Application | Experimental Context |
|---|---|---|
| Racemic γ-Lactam (2-Azabicyclo[2.2.1]hept-5-en-3-one) | Substrate for enantioselective hydrolysis; precursor to (−)-γ-lactam for antivirals [56]. | γ-Lactamase activity and enantioselectivity assays. |
| Chiral HPLC/GC Columns | Analytical separation of enantiomers to determine enantiomeric excess (ee) [57]. | Product analysis in kinetic resolution experiments. |
| Glutaraldehyde | Homobifunctional cross-linker for activating supports with amino groups [11]. | Covalent enzyme immobilization. |
| Chitosan | Natural polymer support for enzyme immobilization; biocompatible and biodegradable [11]. | Carrier for covalent binding or adsorption. |
| Nickel-Nitrilotriacetic Acid (Ni-NTA) Resin | Affinity chromatography medium for purifying recombinant His-tagged proteins [58]. | Purification of heterologously expressed γ-lactamases. |
| Bacillus subtilis Spores | Generally Recognized As Safe (GRAS) platform for enzyme display and immobilization [59]. | Bacterial spore surface display system. |
Navigating Challenges: Optimization Strategies for Extremozyme Production and Deployment
Key Hurdles in Extremophile Cultivation and Biomass Yield
The study of extremophiles—organisms that thrive in conditions once considered incompatible with life—has revolutionized our understanding of biological limits and offers immense potential for biotechnology, medicine, and industrial processes. These remarkable organisms, including thermophiles, psychrophiles, halophiles, acidophiles, and alkaliphiles, produce uniquely stable extremozymes and bioactive compounds that function under harsh conditions where conventional proteins would denature [6] [18]. Their enzymes exhibit exceptional stability—a key property within broader comparative research on microbial versus extremophile enzyme resilience. However, transitioning from laboratory discovery to industrial application presents significant scientific and technical challenges. This review objectively compares the key hurdles in cultivating extremophiles and achieving commercially viable biomass yields, providing researchers with a clear analysis of current limitations and technological solutions. The stringent optimal growth requirements for many extremophiles, combined with the costs associated with their cultivation at scale, impose substantial barriers for industrial applications that must be carefully evaluated against their unique benefits [18] [48].
Major Cultivation Challenges Across Extremophile Types
Extremophile cultivation presents unique, category-specific challenges that impact biomass yield and process economics. The table below systematically compares these hurdles across major extremophile types.
Table 1: Key Cultivation Challenges and Impact on Biomass Yield for Different Extremophile Types
| Extremophile Type | Optimal Growth Conditions | Primary Cultivation Challenges | Impact on Biomass Yield & Economics |
|---|---|---|---|
| Thermophiles (e.g., Thermoanaerobacter sp.) | High temperatures (65-85°C) [60] | • Reduced oxygen solubility at high temperatures [61]• Limited genetic tools for strain improvement• Mixed-product fermentation (e.g., acetate, lactate) reducing ethanol yield to 62-90% of theoretical maximum [60] | • Lower fermentation efficiency in aerobic processes• Suboptimal product yields without extensive metabolic engineering• Wild strains less productive than conventional microbes |
| Halophiles (e.g., Haloferax mediterranei) | High salinity (2-5M NaCl) [61] | • Accelerated corrosion of fermentation equipment [61]• Challenges for process probes and sensors• Residual saline waste after downstream processing• High cost of salts for media preparation | • Increased capital expenditure for specialized equipment• Additional waste treatment requirements• Elevated operational costs for growth media |
| Psychrophiles (e.g., Psychromonas ingrahamii) | Freezing temperatures (-12°C to 10°C) [18] | • High energy demands for temperature maintenance• Slow growth rates and metabolic activity• Specialized harvesting equipment for low-temperature processes | • Lower volumetric productivity• Significant energy input required• Extended process times to achieve target biomass |
| Acidophiles/Alkaliphiles (e.g., Picrophilus oshimae, Bacillus alkaliphilus) | Extreme pH (pH 0-4 or 9-12) [18] [62] | • Specialized corrosion-resistant bioreactors• pH control challenges during scale-up• Limited compatibility with conventional sensors | • Higher capital costs for bioreactor systems• Increased complexity in process control• Potential for process instability during operation |
A critical analysis reveals that these cultivation challenges collectively impact the economic viability of extremophile-based processes. While extremophiles offer advantages like reduced contamination risk in non-sterile conditions [61], the specialized infrastructure requirements and suboptimal yields present significant hurdles. For instance, thermophiles like Thermoanaerobacter sp. produce substantially lower ethanol yields (62-90% of theoretical maximum) compared to conventional Saccharomyces cerevisiae (90-93%) due to their inherent metabolic pathways favoring multiple products [60]. Similarly, the corrosive nature of halophilic cultivation environments accelerates equipment degradation, increasing capital and maintenance costs [61]. These factors must be carefully weighed when considering extremophiles for industrial applications.
Methodological Approaches: Experimental Protocols for Overcoming Cultivation Hurdles
Genetic Tool Development for Extremophile Engineering
Advancing extremophile applications requires sophisticated genetic tools tailored to their unique physiological requirements. The following experimental workflow outlines a generalized protocol for developing genetic manipulation systems for extremophiles, integrating multiple approaches from recent research.
Diagram 1: Genetic Tool Development Workflow for Extremophiles. Key technologies enabling genetic manipulation are highlighted in red.
Protocol Details: Successful implementation requires specialized approaches for different extremophile categories. For thermophiles like Thermus thermophilus, high natural transformation frequencies can be leveraged, though the high G+C content often necessitates gene synthesis for optimal expression [48]. For halophiles, systems have been developed for species including Haloferax mediterranei, leveraging their natural competence or developing shuttle vectors [61]. In methanogenic archaea, specialized techniques account for their anaerobic requirements and unique cellular machinery [48]. The development of inducible promoter systems is particularly critical—for Thermus thermophilus, six inducible promoters (Parg, PdnaK, Pscs-mdh, Pnar, Psip, and PpilA4) have been characterized, though each has limitations (e.g., Pnar requires anaerobic conditions with nitrate, while Psip induction requires silica which inhibits growth) [48]. These tools enable metabolic engineering strategies, such as modifying Thermoanaerobacter strains to reduce byproduct formation and increase ethanol yields from 62-90% to 92-94% of theoretical maximum [60].
Hypotonic Lysis for Biomass Processing from Halophiles
Downstream processing presents significant challenges for extremophile biomass, particularly for intracellular products like polyhydroxyalkanoates (PHA). The following protocol details a method leveraging the unique cellular adaptations of halophiles for efficient product recovery.
Table 2: Experimental Protocol for Hypotonic Lysis-Based Product Recovery from Halophiles
| Step | Procedure | Conditions & Parameters | Mechanistic Basis |
|---|---|---|---|
| 1. Biomass Cultivation | Grow halophilic archaea (e.g., Haloferax mediterranei) in high-salt medium | • 2-4M NaCl concentration• Optimal temperature for target strain• Adequate aeration and nutrient supply | Halophiles accumulate compatible solutes (potassium ions, betaines, ectoines) to maintain osmotic balance in high-salinity environments [61] |
| 2. Biomass Harvesting | Concentrate cells via centrifugation or filtration | • Centrifugation at 8,000-10,000 × g for 15-20 minutes• Maintain elevated salinity during harvesting | Prevents premature cell lysis during processing before the hypotonic shock step |
| 3. Hypotonic Lysis | Resuspend biomass in low-osmotic-strength solution with surfactant | • Distilled water or low-salt buffer• 0.1-0.2% (w/v) Sodium Dodecyl Sulfate (SDS)• Incubation with agitation for 2-4 hours | Osmotic imbalance causes water influx and cell swelling; SDS disrupts lipid membranes, enhancing lysis efficiency [61] |
| 4. Product Recovery | Separate product from cellular debris | • Centrifugation at 12,000 × g for 20 minutes• Wash pellet with appropriate solvents if needed• Dry recovered product | PHA, as intracellular granules, remains in the solid phase while cytoplasmic components solubilize |
| 5. Purity Analysis | Quantify product purity and recovery yield | • Gravimetric analysis• HPLC for polymer composition• Spectroscopy for purity assessment | Target purity: ~95% as demonstrated for both halophilic and thermophilic PHA producers [61] |
Experimental Considerations: This method capitalizes on the fundamental osmoregulation strategies of halophiles. These organisms maintain high intracellular potassium ion concentrations to balance external osmotic pressure. When transferred to hypotonic conditions, water rapidly enters cells, generating tremendous turgor pressure that ruptures the cell envelope [61]. The addition of mild surfactants like SDS enhances membrane disruption without degrading target products. This approach has been successfully applied to both halophiles (Halomonas halophila) and thermophiles (Caldimonas thermodepolymerans), achieving approximately 95% polymer purity while avoiding harsh chemical extraction methods [61]. For acidophiles and alkaliphiles, similar principles can be applied by rapidly shifting pH to induce stress responses that facilitate product release, though specific conditions must be optimized for each species' tolerance range.
Essential Research Tools for Extremophile Cultivation Studies
Advancing extremophile cultivation requires specialized reagents, tools, and platforms. The following table details key solutions for researchers addressing biomass yield challenges.
Table 3: Essential Research Reagent Solutions for Extremophile Cultivation Studies
| Research Tool Category | Specific Examples | Function & Application | Experimental Considerations |
|---|---|---|---|
| Specialized Growth Media | High-salt media for halophiles (2-5M NaCl); Extreme pH buffers for acidophiles/alkaliphiles | Maintains optimal osmotic/ionic conditions for extremophile growth; Prevents cell lysis or stress during cultivation | Cost of specialized components (salts, buffers) significant at scale; May require custom formulation for specific strains [61] |
| Genetic Engineering Tools | CRISPR-Cas systems; Shuttle vectors; Inducible promoter systems (Pnar, Psip for T. thermophilus) | Metabolic engineering to enhance product yields; Gene knockout/knockdown studies; Heterologous expression of biosynthetic pathways | Limited host compatibility; Variable transformation efficiency; Often requires customized development for each extremophile type [48] |
| Extremophile-Specific Bioreactors | Corrosion-resistant alloys for halophile cultivation; High-pressure reactors for piezophiles; Temperature-controlled systems for thermophiles/psychrophiles | Maintains extreme physicochemical conditions during scale-up; Withstands corrosive environments | High capital investment; Specialized maintenance requirements; Limited off-the-shelf availability [61] |
| Metagenomic Tools | Function-based screening of metagenomic libraries; Single-cell genomics; Hi-throughput sequencing | Accessing unculturable extremophile diversity; Identifying novel extremozymes and pathways; Studying community interactions | Expression challenges in heterologous hosts; Limited database annotations for novel sequences [6] [48] |
| Advanced Extraction Methods | Hypotonic lysis solutions; Compatible solute stabilization; Enzyme-stabilizing buffers | Product recovery leveraging extremophile physiology; Maintaining activity of labile extremozymes | Process optimization required for different species; Potential product degradation if not properly stabilized [61] |
These research tools represent critical enabling technologies for overcoming fundamental cultivation challenges. The development of genetic manipulation systems for diverse extremophiles—including thermophiles (Pyrococcus furiosus, Thermococcus spp., Sulfolobus), halophiles, and methanogenic archaea—has been particularly transformative [48]. Similarly, specialized bioreactor designs that accommodate extreme conditions while monitoring key process parameters are essential for reliable scale-up studies. Metagenomic approaches bypass cultivation requirements entirely by directly accessing the genetic potential of extremophile communities, though functional expression of identified genes remains challenging [6].
The cultivation of extremophiles and optimization of their biomass yield present multifaceted challenges spanning genetic, physiological, and process engineering domains. While significant hurdles remain in cost-effective scale-up, yield optimization, and genetic manipulation, continued development of specialized tools and methodologies is steadily addressing these limitations. The unique advantages of extremophiles—including reduced contamination risks, novel catalytic capabilities, and operation under industrially relevant conditions—provide compelling justification for overcoming these barriers. Future research directions should prioritize the expansion of genetic toolkits for underrepresented extremophile groups, integration of omics technologies to guide strain engineering, and development of economical bioreactor systems tailored to extreme conditions. As these advancements mature, extremophiles are poised to play an increasingly significant role in biotechnology, offering sustainable solutions for chemical production, bioremediation, and pharmaceutical applications that leverage their unique stability and catalytic properties.
The pursuit of recombinant enzymes from extremophilic microorganisms presents a significant challenge for biotechnology and drug development. Unlike their mesophilic counterparts, enzymes from extremophiles (extremozymes) possess unparalleled stability under harsh industrial conditions, making them invaluable for processes requiring high temperatures, extreme pH, or the presence of organic solvents [63] [30]. However, their heterologous expression, predominantly in workhorse systems like Escherichia coli, is frequently hampered by low soluble yields, misfolding, and the formation of inactive inclusion bodies [64] [30]. Overcoming these hurdles requires a strategic integration of methods addressing the very foundations of protein synthesis and folding. This guide provides a comparative analysis of three core strategies—codon optimization, chaperone co-expression, and protein refolding—equipping researchers with the data and protocols needed to successfully produce functional extremozymes for their stability research and application development.
Codon Usage Strategies: Optimization vs. Harmonization
The genetic code is degenerate, meaning most amino acids are encoded by multiple synonymous codons. Organisms exhibit a biased usage of these codons, known as codon usage bias, which correlates with the availability of cognate transfer RNAs (tRNAs) to ensure efficient and accurate translation [65] [66]. When expressing a gene from an extremophile in E. coli, a discrepancy between the native codon usage and the host's tRNA pool can cause ribosome stalling, translational errors, and ultimately, protein misfolding [66] [67]. Two primary strategies exist to mitigate this issue: codon optimization and codon harmonization.
Codon optimization involves replacing all rare codons in the native sequence with those that are most frequently used in the expression host. This approach aims to maximize the speed and yield of protein synthesis by aligning translation with the host's preferred codons [67].
Codon harmonization is a more nuanced strategy. It aims to mimic the original rhythm of translation from the native host by adjusting codon usage to preserve regions of slow and fast translation that may be critical for proper co-translational protein folding. Instead of maximizing speed universally, it seeks to replicate the natural translation elongation profile [67].
Table 1: Comparative Analysis of Codon Usage Strategies for Extremozyme Expression.
| Feature | Codon Optimization | Codon Harmonization |
|---|---|---|
| Core Principle | Replace all rare codons with the host's most frequent synonyms [67]. | Mimic the natural translation elongation profile of the native gene [67]. |
| Primary Goal | Maximize translation speed and protein yield [67]. | Promote correct co-translational protein folding [67]. |
| Impact on Soluble Yield | Generally leads to higher levels of soluble protein [67]. | May result in lower soluble yields compared to optimization [67]. |
| Impact on Protein Quality | Can lead to misfolding and reduced specific activity despite high yield [66] [67]. | Often produces proteins with higher thermostability and functional activity [67]. |
| Ideal Use Case | Rapid production of high yields, especially for less complex proteins. | Expression of complex, multi-domain, or folding-sensitive extremozymes. |
Experimental Evidence and Protocol
A direct comparison of these strategies was performed for an N-acetylmuramoyl-L-alanine amidase from the prophage HTH1, identified in the extremophile Hypnocyclicus thermotrophus [67]. The following general workflow can be adapted for any target gene:
Protocol: Comparing Codon Strategies
- Gene Synthesis: Commission the synthesis of the target gene sequence in two variants: one fully optimized for E. coli codon usage and another where codons are harmonized based on the native host's genome.
- Cloning: Clone both gene variants into an appropriate E. coli expression vector (e.g., pET series).
- Expression: Transform the plasmids into a suitable E. coli strain (e.g., BL21(DE3)). Induce expression with IPTG under standardized conditions (e.g., OD600, temperature, induction time).
- Analysis:
- Soluble Yield: Lyse the cells, separate soluble and insoluble fractions via centrifugation, and analyze by SDS-PAGE with densitometry or quantify via Western blot.
- Thermostability: Purify the soluble proteins and use a technique like differential scanning fluorimetry (thermal shift assay) to determine the melting temperature (Tm). Incubate aliquots at a defined elevated temperature (e.g., 60°C) for a set time, then measure residual activity.
Result Interpretation: In the cited study [67], the optimized variant yielded more soluble protein, but the harmonized version demonstrated superior thermostability, retaining a higher level of activity after heat challenge. This underscores that for extremozymes where stability is the key property, codon harmonization, despite potentially lower yields, may be the preferred strategy.
Chaperone Co-expression for In Vivo Folding
Molecular chaperones are host proteins that assist in the folding, assembly, and stabilization of other proteins. Co-expressing recombinant chaperones alongside the target extremozyme can significantly enhance the solubility and functional yield by preventing aggregation and facilitating proper folding within the cell [30].
Table 2: Key Chaperone Systems for Co-expression in E. coli.
| Chaperone System | Primary Function | Role in Extremozyme Expression |
|---|---|---|
| GroEL/GroES (Hsp60/Hsp10) | Forms a barrel-like complex that provides an isolated chamber for single polypeptide chains to fold [68]. | Essential for folding many proteins; co-expression is crucial for complex extremozymes prone to misfolding [30]. |
| DnaK/DnaJ/GrpE (Hsp70 System) | Binds to hydrophobic patches of nascent chains, preventing aggregation and facilitating folding [68]. | Ideal for preventing initial misfolding of thermophilic enzymes being produced in a mesophilic host [30]. |
| Trigger Factor | A ribosome-associated chaperone that interacts with nascent chains co-translationally [68]. | Provides the first line of defense against misfolding; can be co-expressed to enhance solubility [30]. |
| Small Heat-Shock Proteins (sHsps) | Act as holdases, binding unfolded proteins and preventing aggregation under stress conditions [68]. | Particularly useful when expressing psychrophilic enzymes in E. coli, as the higher temperature may induce stress [30]. |
Experimental Protocol for Chaperone Co-expression
Protocol: Screening Chaperone Plasmid Kits
- Selection: Obtain a commercial chaperone plasmid set (e.g., Takara's Chaperone Plasmid Set), which typically includes plasmids encoding various combinations of GroEL/GroES, DnaK/DnaJ/GrpE, Trigger Factor, and others.
- Co-transformation: Co-transform the target extremozyme expression plasmid with individual or combined chaperone plasmids into the expression host. Include a control with an empty plasmid.
- Dual Selection: Grow cultures in media containing antibiotics for both plasmids.
- Expression and Analysis: Induce both the target gene and the chaperone genes according to the specific protocols for each system (e.g., chaperone expression may require a separate inducer like L-arabinose). Analyze cell lysates for soluble target protein using SDS-PAGE and functional activity assays.
The workflow below illustrates the decision-making process for selecting and implementing a chaperone co-expression strategy.
Diagram 1: Chaperone co-expression strategy selection.
Recovery of Bioactive Protein from Inclusion Bodies
Despite the best preventive strategies, the high-level expression of extremozymes in E. coli often results in the formation of inclusion bodies—densely packed aggregates of misfolded protein [64]. While traditionally viewed as a setback, inclusion bodies can also be a source of highly pure protein, requiring a robust refolding protocol.
Mild Solubilization and Refolding Protocol
Classical methods use high concentrations of chaotropes like urea or guanidine-HCl, which fully denature the protein and can complicate refolding. "Mild solubilization" leverages the finding that proteins in inclusion bodies can retain native-like secondary structure, enabling solubilization with lower denaturant concentrations and preserving functionality [64].
Protocol: Mild Solubilization and Refolding [64]
- Isolation and Washing: Harvest bacterial cells by centrifugation. Lyse using a French press or sonication. Centrifuge at high speed (e.g., 15,000 x g) to pellet inclusion bodies. Wash the pellet repeatedly with buffer containing Triton X-100 and EDTA to remove membrane and nucleic acid contaminants.
- Mild Solubilization: Solubilize the inclusion body pellet in a buffer containing a low concentration of a chaotropic agent (e.g., 2-4 M urea) or a denaturing surfactant (e.g., N-Lauroylsarcosine). The use of alkaline pH (e.g., pH 9.0-10.5) can also be effective for some proteins. The key is to use the minimal denaturant strength required for solubilization.
- Refolding: Dilute the solubilized protein rapidly into a large volume (typically 50-100 fold) of refolding buffer. This buffer is critical and should be optimized for the specific protein. Key components include:
- Redox System: A pair of low-molecular-weight thiols like reduced/oxidized glutathione (GSH/GSSG) or cysteine/cystamine to facilitate disulfide bond reshuffling.
- Additives: L-arginine (0.5-1 M) is highly effective at suppressing aggregation. Polyethylene glycol (PEG) and glycerol can also enhance correct folding.
- Stabilizing Ions: Ca²⁺ or other cofactors required for the extremozyme's activity.
- Concentration and Purification: Concentrate the refolded protein using tangential flow filtration or centrifugal concentrators. Purify the active fraction using size-exclusion or ion-exchange chromatography to separate correctly folded monomers from aggregates or misfolded species.
The following workflow visualizes the key stages of this process.
Diagram 2: Protein recovery from inclusion bodies.
The Scientist's Toolkit: Essential Reagents and Solutions
Table 3: Key Research Reagent Solutions for Heterologous Expression.
| Reagent/Solution | Function | Example Use Case |
|---|---|---|
| Codon-Optimized/Harmonized Genes | Gene synthesis services provide sequences tailored for expression in E. coli, bypassing native codon bias issues [67]. | Essential first step for expressing any extremozyme gene from archaea or other GC-rich organisms. |
| Commercial Chaperone Plasmid Kits | Pre-configured plasmids for co-expressing various E. coli chaperone systems (e.g., GroEL/S, DnaK/J/E) [30]. | Systematic screening for the best chaperone system to improve soluble yield of a target extremozyme. |
| L-Arginine Hydrochloride | An additive in refolding buffers that suppresses protein aggregation by non-specifically interacting with folding intermediates [64]. | Added to 0.5-1 M in refolding buffers to increase the yield of correctly folded protein during dilution. |
| Reduced/Oxidized Glutathione (GSH/GSSG) | A redox couple that creates a buffer system for disulfide bond formation and reshuffling in the oxidative folding of proteins [64]. | Critical for refolding extremozymes that require disulfide bonds for their stability and activity. |
| N-Lauroylsarcosine | A mild, denaturing surfactant effective at solubilizing inclusion bodies while often preserving native-like structure better than strong chaotropes [64]. | Used as an alternative to urea for the mild solubilization step of inclusion body processing. |
No single strategy is universally superior. Success in expressing a functional extremozyme often requires an iterative and integrated approach. A recommended workflow begins with testing both codon-optimized and codon-harmonized gene constructs. If soluble yield remains low, chaperone co-expression should be screened. The formation of inclusion bodies, while a setback, is not a dead end, as robust refolding protocols can recover active protein.
The choice of strategy is deeply intertwined with the broader research on comparative enzyme stability. The expression and folding efficiency of an enzyme is a direct reflection of its innate structural adaptability. By systematically comparing the success of these optimization strategies for mesophilic versus extremophilic enzymes, researchers can glean fundamental insights into the folding landscapes and evolutionary constraints that define protein stability. The experimental data and protocols provided here serve as a foundational guide for this ongoing exploration, enabling the successful production of these valuable biocatalysts.
The unique structural and functional adaptations of extremozymes—enzymes derived from organisms thriving in extreme environments—make them invaluable biocatalysts for industrial processes, including pharmaceutical manufacturing [18]. These enzymes exhibit exceptional stability and functionality under harsh conditions, such as extreme temperatures, pH, and salinity, that would denature most conventional enzymes [63]. However, naturally occurring extremozymes often require further optimization to meet the specific demands of industrial applications, driving the development of advanced protein engineering strategies. Within the broader context of comparative stability research between microbial and extremophile enzymes, two powerful engineering paradigms have emerged: rational design and directed evolution [30]. This guide provides a comparative analysis of these methodologies, offering experimental data and protocols to inform selection for biocatalyst development projects.
Fundamental Principles and Comparative Analysis
Rational design employs knowledge of protein structure, function, and mechanism to make targeted mutations through site-directed mutagenesis. This approach requires detailed structural information and computational analysis to predict amino acid substitutions that will enhance desired properties [69]. In contrast, directed evolution mimics natural selection in laboratory settings through iterative rounds of random mutagenesis and screening to identify variants with improved traits, without requiring prior structural knowledge [30]. A third hybrid approach, semi-rational design, combines elements of both strategies by using structural and evolutionary information to create focused, "smart" libraries targeting specific protein regions, thereby reducing the screening burden [69].
Table 1: Core Methodology Comparison: Rational Design vs. Directed Evolution
| Aspect | Rational Design | Directed Evolution | Semi-Rational Design |
|---|---|---|---|
| Knowledge Requirement | High (3D structure, mechanism) | Low | Medium (sequence, homology) |
| Library Size | Small (tens to hundreds) | Very large (thousands to millions) | Focused (hundreds to thousands) |
| Technical Emphasis | Computational prediction, targeted mutagenesis | Random diversity generation, high-throughput screening | Target selection, restricted amino acid sets |
| Primary Advantage | Precise, minimal mutations | No structural information needed | Balanced efficiency and coverage |
| Key Limitation | Limited to predictable improvements | Extensive screening required | Requires some evolutionary information |
Experimental Protocols and Workflows
Workflow Visualization
Key Experimental Protocols
Rational Design: Structure-Guided Engineering
The strategic engineering of a thermophilic lipase from Thermomicrobium roseum (TrLipE) demonstrates rational design principles. Researchers created 18 chimeras through lid swapping—replacing structural elements that control access to the active site. Molecular dynamics simulations revealed increased flexibility in these variants, which exhibited enhanced thermostability and broader pH tolerance compared to the wild-type enzyme and other thermostable lipases. Specific variants with single, double, or triple substitutions demonstrated 2-3-fold faster catalysis than the wild-type enzyme [70].
Detailed Protocol: Site-Directed Mutagenesis for Targeted Improvements
- Structural Analysis: Obtain 3D structure via X-ray crystallography or cryo-electron microscopy [70]
- Target Identification: Use computational tools to identify residues critical for stability, specificity, or activity
- Primer Design: Design mutagenic primers with 15-20 bp flanking the mutation site
- PCR Amplification: Perform PCR with high-fidelity DNA polymerase
- Template Digestion: Digest methylated template DNA with DpnI enzyme
- Transformation: Introduce plasmid into competent E. coli cells
- Screening: Verify mutations by DNA sequencing
- Characterization: Express and purify variant enzymes for biochemical analysis
Directed Evolution: Diversity-Driven Optimization
A study addressing the challenge of low yields in recombinant extremozyme production exemplifies directed evolution. When expressing extremozymes in conventional mesophilic hosts like E. coli, the enzymes often form inclusion bodies or show poor solubility. Directed evolution approaches target the enzyme sequence itself to improve compatibility with heterologous expression systems while maintaining extremophilic properties [30].
Detailed Protocol: Iterative Random Mutagenesis and Screening
- Diversity Generation: Use error-prone PCR or DNA shuffling to create mutant libraries
- Library Construction: Clone variants into expression vectors
- High-Throughput Expression: Express variants in multi-well plates
- Activity Screening: Implement robotic screening under desired conditions (e.g., high temperature, extreme pH)
- Hit Identification: Select top performers based on catalytic efficiency and stability
- Gene Recombination: Use staggered extension process or DNA shuffling to combine beneficial mutations
- Iterative Cycles: Repeat process for 3-5 generations to accumulate improvements
Performance Data and Comparative Stability
Quantitative Comparison of Engineering Outcomes
Table 2: Experimental Performance Data from Engineered Extremozymes
| Enzyme / Source | Engineering Method | Key Mutations/Approach | Catalytic Efficiency Improvement | Stability Enhancement |
|---|---|---|---|---|
| TrLipE lipase (Thermomicrobium roseum) | Semi-rational | Lid swapping chimera creation | 2-3-fold increase in catalytic rate [70] | Enhanced thermostability & broader pH range [70] |
| α-amylase (Geobacillus stearothermophilus) | Rational | Mutation of "hinge" positions in motion pathway | 93.8% increase in catalytic efficiency [70] | Optimal temperature of 90°C maintained [70] |
| SZ_A polymerase (Salt-tolerant variant) | Semi-rational | Conserved amino acid substitution | Improved salt tolerance & processivity [70] | Exonuclease deficiency for nanopore sequencing [70] |
| Uricase TrUox (Thermoactinospora rubra) | Natural extremozyme | None (native enzyme) | High catalytic efficiency at neutral pH | Maintained activity after 4 days at 50°C [71] |
Stability Metrics in Comparative Context
When evaluating engineered extremozymes against their mesophilic counterparts, several key stability parameters demonstrate the advantage of starting with extremophilic scaffolds:
Thermal Stability: Engineered thermophilic enzymes typically maintain folded structures and activity at temperatures 20-40°C higher than mesophilic equivalents. For instance, uricase TrUox from Thermoactinospora rubra maintained activity after 4 days at 50°C, a stability profile that would require extensive engineering in mesophilic enzymes [71].
Solvent Tolerance: Extremozymes engineered through both rational and evolutionary approaches show enhanced resistance to organic solvents, with some variants maintaining activity in up to 50% cosolvent concentrations. This is particularly valuable for pharmaceutical synthesis where substrate solubility often requires mixed aqueous-organic systems [30].
pH Robustness: Engineering can further expand the already broad pH tolerance of native extremozymes. The engineered TrLipE variants displayed improved activity across a wider pH range compared to other thermostable lipases [70].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents for Extremozyme Engineering
| Reagent / Material | Function | Example Applications |
|---|---|---|
| Error-Prone PCR Kits | Generate random mutations for directed evolution | Creating diverse mutant libraries [30] |
| Site-Directed Mutagenesis Kits | Introduce specific, targeted mutations | Rational design of active site residues [69] |
| Thermostable DNA Polymerases | PCR amplification of extremozyme genes | Amplifying GC-rich templates from thermophiles [70] |
| Chitosan & Other Natural Polymers | Enzyme immobilization matrices | Stabilizing engineered enzymes for reuse [11] |
| Bacterial Spore Display Systems | Enzyme stabilization via surface display | Enhancing operational stability under harsh conditions [59] |
| Computational Design Software | Predict effects of amino acid substitutions | Rational design and semi-rational library design [69] |
The choice between rational design and directed evolution for extremozyme engineering depends on multiple project-specific factors. Rational design offers precision and efficiency when structural information is available and targeted improvements are sought. The lid-swapped TrLipE lipase variants exemplify how structural insights can guide specific enhancements in catalytic efficiency and stability [70]. Conversely, directed evolution provides a powerful discovery platform when structural data is limited or when seeking unexpected solutions to complex engineering challenges, such as improving heterologous expression of extremozymes in mesophilic hosts [30].
The emerging trend of semi-rational design represents a synergistic approach, leveraging evolutionary information and computational power to create focused libraries that maximize functional diversity while minimizing screening requirements [69]. As structural databases expand and computational tools become more sophisticated, this integrated approach promises to accelerate the development of next-generation extremozymes for pharmaceutical applications and beyond, ultimately enhancing the comparative stability advantages of extremophile-derived enzymes over their mesophilic counterparts.
Immobilization Techniques to Boost Operational Stability and Reusability
Enzymes, as biological catalysts, are indispensable in modern biotechnology, finding applications across diverse sectors including pharmaceuticals, food processing, and bioenergy [72]. Their specificity, efficiency, and ability to operate under mild conditions make them superior to traditional chemical catalysts. However, the widespread industrial application of free enzymes is often hampered by inherent limitations such as poor stability under operational conditions (e.g., elevated temperatures, extreme pH), short shelf life, and the inability to be reused, which significantly increases process costs [72] [73]. Enzyme immobilization, the process of confining or localizing enzymes to a solid phase distinct from the substrate and product phase, has emerged as a powerful strategy to overcome these challenges [74].
Immobilization enhances enzyme applicability by improving stability, facilitating easy recovery and reuse, and protecting the enzyme from harsh environmental conditions [72] [75]. This guide provides a comparative analysis of major immobilization techniques, supported by experimental data, and frames the discussion within the broader context of stabilizing enzymes from both conventional microbial sources and robust extremophiles.
Comparison of Core Immobilization Techniques
The selection of an immobilization technique is crucial as it directly impacts the enzyme's performance, stability, and cost-effectiveness. The following sections and comparative table detail the most common methods.
Table 1: Comparison of Key Enzyme Immobilization Techniques
| Technique | Mechanism of Binding | Advantages | Disadvantages | Best Suited For |
|---|---|---|---|---|
| Adsorption [74] [73] [76] | Weak forces (van der Waals, ionic, hydrophobic, hydrogen bonding) | Simple, reversible, low cost, minimal enzyme structure alteration [76]. | Enzyme leakage (desorption) due to weak bonds, leading to product contamination [73]. | Rapid, cost-sensitive processes where mild conditions can be maintained. |
| Covalent Binding [74] [73] [77] | Strong, irreversible covalent bonds between enzyme and support | No enzyme leakage, high stability, easy substrate contact, enhanced thermal stability [73] [77]. | Harsh conditions can reduce activity; potential denaturation; relatively expensive supports [73]. | Processes requiring long-term operational stability and no enzyme contamination. |
| Entrapment/ Encapsulation [74] [73] | Physical confinement within a porous polymer matrix or membrane | Enzyme protected from harsh environments and microbial contamination [74]. | Diffusion limitations, reduced reaction rates, possible enzyme leakage from pores [74] [73]. | Enzymes with small substrates where diffusion is not a major constraint. |
| Cross-linking [74] [73] | Enzyme molecules linked to each other via multifunctional reagents (e.g., glutaraldehyde) | High stability, no separate support needed, reusable aggregates [74]. | Can be costly to optimize, may reduce enzyme activity, mass transfer limitations [74] [76]. | Stabilizing enzyme aggregates when support-free operation is desired. |
The following decision pathway visualizes the process of selecting an appropriate immobilization technique based on key criteria:
Experimental Protocols & Performance Data
Case Study: Covalent Immobilization of β-Glucosidase
A detailed study on the covalent immobilization of a thermostable β-glucosidase from Thermotoga maritima onto sodium alginate-based magnetic nanoparticles (SA-Fe₃O₄ MNPs) provides robust experimental data on performance enhancement [78].
Experimental Protocol:
- Support Synthesis & Activation: SA-Fe₃O₄ MNPs were synthesized and activated using a cross-linker (e.g., glutaraldehyde), creating electrophilic groups on the carrier surface [73] [78].
- Enzyme Binding: The β-glucosidase enzyme was covalently coupled to the activated support via its nucleophilic amino acid residues (e.g., lysine). The interaction was confirmed using SDS-PAGE, TEM, and FT-IR spectroscopy [78].
- Characterization: The kinetic parameters (Km, Vmax), optimum pH and temperature, thermal stability, and reusability of the immobilized enzyme were systematically compared to its free form [78].
Table 2: Experimental Performance Data of Free vs. Immobilized β-Glucosidase [78]
| Parameter | Free Enzyme | Immobilized Enzyme | Change |
|---|---|---|---|
| Optimum Temperature | Not specified | 85 °C | Increased |
| Optimum pH | Not specified | 7.0 | Maintained (neutral) |
| Kinetic Parameter (Km) | Baseline | Increased by 65% | Slightly reduced affinity |
| Kinetic Parameter (Vmax) | Baseline | Increased by 41% | Higher reaction rate |
| Thermal Stability | Baseline | Enhanced | Higher resistance to denaturation |
| Operational Stability | Single use | ~40% activity retained after 6 reuse cycles | Excellent reusability |
Case Study: Adsorption-Based Immobilization for Reusability
Another experiment illustrates the application and limitations of adsorption-based methods in a real-world fermentation process.
Experimental Protocol:
- Cell Adsorption: A consortium of yeast and acetic acid bacteria was immobilized via adsorption onto natural supports like corncobs (CC) and oakwood chips (OWC) by submerging the supports in the microbial culture [79].
- Fermentation & Reuse: The immobilized cells were used for balsamic-styled vinegar production. After the first fermentation cycle, the supports (with adsorbed cells) were recovered, rinsed, and inoculated into a fresh medium for a second cycle [79].
- Performance Analysis: Microbial activity and product formation rates were monitored for both cycles. Scanning Electron Microscopy (SEM) was used to evaluate the structural integrity of the supports after use [79].
Findings: The study found a substantial decline in microbial activity and acetification rates when the adsorbed cells were reused for the second cycle. This was attributed to cell desorption and a loss of viable cells from the supports, highlighting a key drawback of the adsorption method under operational conditions [79].
The Stability Frontier: Microbial vs. Extremophile Enzymes
A core thesis in modern enzymology is the comparative stability of enzymes derived from mesophilic microorganisms versus those from extremophiles—organisms that thrive in extreme habitats [14].
- Microbial Enzymes (Mesophiles): Enzymes from conventional sources typically operate under mild conditions. Their stability often needs to be engineered post-harvest, primarily through the immobilization techniques discussed herein, to make them suitable for industrial processes [72] [73].
- Extremophile Enzymes (Extremozymes): Extremophiles produce naturally robust extremozymes that are intrinsically stable under harsh conditions [14] [30]. For instance:
- Thermozymes from thermophiles exhibit superior stability at high temperatures due to structural adaptations like a high content of charged amino acids, hydrophobic interactions, and disulfide bonds [14].
- Psychrophilic Enzymes from cold-adapted organisms possess high molecular flexibility to function efficiently at low temperatures, achieved through reduced proline and arginine content and fewer hydrogen bonds [14].
The following diagram illustrates the complementary strategies of using extremophile enzymes and immobilization to achieve operational stability.
Immobilization can further augment the innate stability of extremozymes. Research shows that immobilizing a thermostable β-glucosidase from Thermotoga maritima not only allowed easy recycling but also raised its optimum temperature to 85°C [78], demonstrating a synergistic effect between intrinsic and engineered stability.
The Scientist's Toolkit: Essential Research Reagents
The following table details key reagents and materials essential for conducting immobilization experiments, drawing from the protocols cited.
Table 3: Key Research Reagent Solutions for Enzyme Immobilization
| Reagent/Material | Function/Application | Examples from Literature |
|---|---|---|
| Support Matrices | Provides a solid surface or matrix for enzyme attachment. | Sodium alginate (entrapment) [78], Octyl-agarose (adsorption) [74], Chitosan (covalent/adsorption) [73], Mesoporous Silica Nanoparticles (MSNs) [74] [73], Magnetic Nanoparticles (Fe₃O₄) [78]. |
| Activation Reagents/Cross-linkers | Activates support surfaces or creates covalent bonds between enzyme and support or between enzyme molecules. | Glutaraldehyde [74] [73], Carbodiimide [73] [77], Bifunctional reagents (e.g., bisdiazobenzidine) [74]. |
| Characterization Tools | Used to confirm and analyze the success of the immobilization process. | SDS-PAGE, FT-IR Spectroscopy, Transmission Electron Microscope (TEM) [78], Scanning Electron Microscope (SEM) for structural integrity [79]. |
The strategic application of immobilization techniques is fundamental to enhancing the operational stability and reusability of enzymes, thereby making biocatalytic processes more industrially viable and cost-effective. As summarized in this guide, the choice between methods like adsorption, covalent binding, entrapment, and cross-linking involves a careful trade-off between stability, cost, simplicity, and retained activity.
The ongoing research in this field is increasingly focused on hybrid approaches, such as using novel support materials like magnetic nanoparticles and combining immobilization with the innate robustness of extremozymes. This powerful synergy promises to unlock new possibilities in biotechnology, enabling more efficient and sustainable industrial processes in drug development, bioenergy, and beyond.
Scaling up enzyme production from laboratory-scale bioreactors to industrial-scale manufacturing presents a complex set of engineering and biological challenges. This process involves navigating the delicate balance between maintaining optimal conditions for enzyme production and adapting to the physical realities of large-scale equipment. For researchers comparing microbial and extremophile enzymes, understanding these scale-up principles is crucial, as the unique stability properties of extremophile-derived enzymes can significantly influence scaling strategy success.
The fundamental obstacle in scale-up arises from nonlinear changes in physical parameters as bioreactor volume increases. While small-scale bioreactors offer homogeneous conditions, industrial-scale vessels frequently develop gradients in temperature, dissolved oxygen, pH, and substrate concentration [80]. These gradients can alter cellular physiology, ultimately affecting both the yield and catalytic properties of the produced enzymes. The scaling process typically progresses from high-throughput miniaturized bioreactors (15–250 mL) through bench-scale (1–10 L) to pilot and production-scale bioreactors (200–5,000 L or more) [80].
This guide objectively compares scaling methodologies for conventional microbial enzymes versus those derived from extremophiles, with a focus on how their inherent stabilities impact process design and economic viability.
Comparative Analysis: Microbial vs. Extremophile Enzymes
Fundamental Stability Differences
Enzymes derived from extremophiles possess intrinsic structural advantages that translate directly to improved performance under industrial conditions. Thermophilic enzymes from organisms adapted to high temperatures (60–110°C) exhibit enhanced stability through multiple molecular mechanisms, including increased charged surface residues (Glu, Arg, Lys), additional salt bridges, strengthened hydrophobic cores, and shortened surface loops [2]. These adaptations result in superior resistance to the thermal and chemical stresses encountered in industrial processes.
In contrast, psychrophilic enzymes from cold-adapted organisms feature strategic flexibility achieved through reduced hydrophobic interactions, decreased proline content, and fewer ion pairs, making them highly efficient at lower temperatures and potentially reducing energy costs for processes requiring moderate temperatures [2]. Halophilic enzymes from high-salt environments maintain stability in low-water conditions through abundant negatively charged surface residues, a valuable trait for applications in non-aqueous systems [9].
Industrial Process Implications
The stability advantages of extremozymes directly impact scaling economics by reducing the need for precise environmental control and extending functional lifespan. Conventional microbial enzymes often require stricter parameter control during both production and catalysis, increasing operational complexity and cost at industrial scale [39].
Table 1: Comparative Enzyme Stability Under Industrial Conditions
| Enzyme Characteristic | Conventional Microbial | Extremophile-Derived | Industrial Impact |
|---|---|---|---|
| Thermal Stability | Moderate (30-60°C) | High (60-110°C for thermophiles) | Reduced cooling requirements, higher process temperatures |
| pH Tolerance | Narrow range (typically pH 5-8) | Broad range (pH <2->9 for acid/alkaliphiles) | Reduced pH control infrastructure |
| Organic Solvent Stability | Generally low | Moderate to high (especially halophiles) | Enables non-aqueous biocatalysis |
| Operational Half-life | Hours to days | Days to weeks | Reduced enzyme replenishment costs |
| Protease Resistance | Variable | Often enhanced | Improved longevity in complex mixtures |
Experimental Data and Performance Comparison
Quantitative Stability Metrics
Rigorous laboratory testing provides essential data for predicting scale-up performance. The following experimental protocols enable researchers to generate comparative stability profiles for enzyme candidates.
Protocol 1: Thermal Stability Assessment
- Preparation: Purify enzyme samples to >95% homogeneity via affinity chromatography.
- Heat Challenge: Incalate separate aliquots at temperatures ranging from 30°C to 100°C in a thermal cycler or water baths with temperature monitoring.
- Sampling: Withdraw samples at predetermined time points (0, 15, 30, 60, 120 minutes).
- Activity Assay: Measure residual activity using standard assays (e.g., spectrophotometric substrate conversion).
- Data Analysis: Calculate half-life at each temperature and determine melting temperature (Tm) via differential scanning calorimetry.
Protocol 2: pH Stability Profile
- Buffer Series: Prepare buffers across pH 2.0-10.0 with appropriate buffering capacity.
- Incubation: Incubate enzymes in each buffer for 24 hours at 25°C.
- Measurement: Assess residual activity under standard assay conditions.
- Analysis: Plot relative activity versus pH to determine optimal range and stability boundaries.
Table 2: Experimental Stability Data for Representative Enzymes
| Enzyme Source | Optimal Temp (°C) | Thermal Half-life | pH Range | Solvent Tolerance |
|---|---|---|---|---|
| E. coli Mesophilic Protease | 45 | 45 min at 50°C | 6.0-8.0 | <15% organic cosolvent |
| Thermus aquaticus Taq Polymerase | 72 | 40 min at 95°C | 7.0-9.0 | Moderate |
| Pyrococcus furiosus Glutamate Dehydrogenase | 85 | 8 h at 90°C | 6.0-8.5 | High |
| Bacillus halodurans Xylanase | 65 | 3 h at 70°C | 6.0-10.0 | Moderate |
| Sulfolobus solfataricus γ-Lactamase | 85 | 6 h at 80°C (immobilized) | 5.0-9.0 | High |
Experimental data demonstrates that extremophile-derived enzymes typically maintain significant activity under conditions that completely inactivate their mesophilic counterparts. For example, the triple-point mutant R77F/E145M/T284R of a thermophilic xylanase exhibited a 3.39-fold increase in specific activity while simultaneously increasing melting temperature by 2.4°C, overcoming the traditional stability-activity trade-off [81].
Advanced Engineering Strategies
Machine learning approaches now enable more sophisticated enzyme engineering for industrial applications. The iCASE (isothermal compressibility-assisted dynamic squeezing index perturbation engineering) strategy creates hierarchical modular networks to optimize enzymes of varying complexity [81]. This method identifies key regulatory residues outside active sites that influence both stability and activity, allowing simultaneous improvement of both properties.
Diagram 1: Enzyme Scaling Workflow
Scaling Methodologies and Practical Implementation
Bioreactor Scale-Up Principles
Successful scale-up requires careful management of both scale-independent and scale-dependent parameters. Scale-independent parameters including pH, temperature, dissolved oxygen concentration, and media composition can be optimized at small scale and maintained constant during scale-up [80]. Scale-dependent parameters such as impeller rotational speed, gas-sparging rates, and working volume must be systematically adjusted to accommodate changing hydrodynamic conditions [80].
The most common scaling criteria include:
- Constant Power per Unit Volume (P/V): Maintains similar mixing energy but increases shear stress and tip speed.
- Constant Oxygen Mass Transfer Coefficient (kLa): Ensures consistent oxygen delivery but may dramatically increase power input.
- Constant Impeller Tip Speed: Limits shear damage to cells but reduces mixing efficiency.
- Constant Mixing Time: Difficult to maintain as it requires disproportionately high power input.
Table 3: Scale-Up Parameter Interdependence (Scale Factor: 125x)
| Scale-Up Criterion | Power/Volume | Tip Speed | Mixing Time | kLa | Reynolds Number |
|---|---|---|---|---|---|
| Equal P/V | 1 | 2.2 | 2.7 | 1.6 | 11.2 |
| Equal Tip Speed | 0.2 | 1 | 5 | 0.5 | 5 |
| Equal kLa | 1.6 | 2.7 | 2.2 | 1 | 14.7 |
| Equal Mixing Time | 25 | 5 | 1 | 7.5 | 25 |
Practical Scaling Challenges and Solutions
Mixing and Gradient Formation: As bioreactor size increases, mixing time increases significantly, leading to substrate, pH, and oxygen gradients. In large-scale cell-culture bioreactors, mixing times can reach several minutes [80]. Cells circulating through these gradients experience a constantly changing environment that can alter metabolism and enzyme production.
Heat Transfer Limitations: The surface-area-to-volume ratio decreases dramatically with scale, creating challenges for heat removal, particularly in high-density microbial fermentations [80].
Gas Transfer Efficiency: Oxygen transfer and carbon dioxide stripping become increasingly difficult at larger scales due to increased hydrostatic pressure and reduced surface area at the bioreactor headspace [80].
Diagram 2: Bioreactor Scaling Strategy
The Scientist's Toolkit: Research Reagent Solutions
Table 4: Essential Research Tools for Enzyme Scaling Studies
| Reagent/Equipment | Function | Application Note |
|---|---|---|
| Lab-Scale Bioreactors (1-10L) | Process parameter optimization | Maintain geometric similarity to production scale |
| Stability Screening Assays | Thermal, pH, and solvent stability profiling | Use high-throughput formats for rapid candidate screening |
| Metagenomic Libraries | Novel enzyme discovery from extreme environments | Function-based screening identifies novel activities [9] |
| Machine Learning Platforms | Predictive enzyme engineering | iCASE strategy balances stability-activity trade-off [81] |
| Immobilization Supports | Enzyme stabilization and reusability | Cross-linked enzyme preparations enhance thermostability [39] |
| Advanced Analytics | Product quality assessment | HPLC, MS, and activity assays monitor consistency across scales |
Scaling enzyme production from laboratory to industrial scale requires a methodical approach that acknowledges both engineering constraints and biological realities. Enzymes derived from extremophiles offer distinct advantages in scaling due to their inherent stability under process-relevant conditions. By integrating modern tools like machine learning-guided engineering with traditional scale-up principles, researchers can develop more robust and economically viable enzyme production processes.
The future of industrial enzyme production will increasingly leverage extremophile diversity discovered through metagenomic mining and enhanced through rational design. These advances will help overcome the current challenges in scaling and enable more sustainable industrial processes across pharmaceutical, energy, and chemical sectors.
Performance Under Pressure: Direct Comparative Analysis of Enzyme Stability
The comparative analysis of enzyme stability, particularly between microbial and extremophile-derived enzymes, is a cornerstone of modern enzymology and biocatalyst development. For researchers and drug development professionals, selecting the appropriate assessment methodology is critical, as the data directly informs decisions on enzyme selection for industrial processes, therapeutic applications, and further protein engineering efforts [63] [39]. Stability dictates an enzyme's functional lifetime, reaction efficiency, and economic viability in applications ranging from pharmaceutical synthesis to diagnostic assays [9] [82].
This guide provides a comparative analysis of the predominant experimental and computational methodologies used to quantify enzyme stability. It evaluates these techniques based on key performance metrics, including throughput, information depth, and relevance to real-world operational conditions, thereby offering a framework for selecting optimal assessment strategies for specific research and development goals.
Key Stability Metrics and Their Significance
The stability of an enzyme is quantified through several key metrics, each providing unique insight into its structural robustness. The choice of metric often depends on the denaturing agent used in the assay.
- Thermodynamic Stability (ΔG, ΔΔG): This represents the free energy difference between the folded (native) and unfolded (denatured) states of the enzyme. A more positive ΔG indicates greater intrinsic stability. The change in free energy upon mutation (ΔΔG) is a fundamental parameter for understanding the impact of protein engineering [83] [84].
- Melting Temperature (Tₘ): The temperature at which 50% of the enzyme population is unfolded. A higher Tₘ denotes greater thermal stability and is often used as a rapid comparative screen [83].
- Denaturation Midpoint (Cₘ or D₁/₂): The concentration of a chemical denaturant (e.g., guanidinium chloride or urea) required to unfold 50% of the enzyme population. This metric reflects the enzyme's resistance to chemical denaturation [83].
- Half-life (t₁/₂): The time required for an enzyme to lose 50% of its initial activity under defined conditions (e.g., at a specific temperature or pH). This is a critical practical metric for industrial applications, directly relating to operational longevity [39].
Comparative Analysis of Experimental Assays
The following table summarizes the core experimental techniques used for assessing enzyme stability, comparing their primary applications, advantages, and limitations.
Table 1: Comparison of Key Experimental Assays for Enzyme Stability Assessment
| Method | Measured Metric(s) | Throughput | Key Advantage(s) | Key Limitation(s) |
|---|---|---|---|---|
| Differential Scanning Calorimetry (DSC) | Tₘ, ΔH (enthalpy) | Low | Directly measures heat capacity changes; provides thermodynamic parameters. | Low throughput; requires high protein concentrations; may miss non-cooperative unfolding [85]. |
| Spectroscopic Denaturation (e.g., Fluorescence, CD) | Cₘ, Tₘ, ΔG, m-value | Medium | Probes structural integrity; can be performed on plate readers for higher throughput. | Requires a spectroscopic signal (e.g., Trp, secondary structure); data analysis assumes a two-state model [83] [85]. |
| Plate-Based Activity/Stability Screen | Residual activity after stress | High | Functional readout; can screen thousands of variants (e.g., for directed evolution). | Often indirect stability measure; conditions may not be at equilibrium [9] [86]. |
| Live Bacterial Cell Assay (LEICA) | Microbial growth rate | High | In vivo functional context; high-throughput screening of mutant libraries. | Limited to enzymes that can complement microbial metabolism; growth rate is an indirect proxy [86]. |
Detailed Experimental Protocols
Plate-Based Chemical Denaturation with Fluorescence Detection
This medium- to high-throughput protocol is adapted from automated stability screening pipelines and is suitable for characterizing both microbial and extremophile enzymes [83].
Workflow Overview:
Key Steps:
- Protein Purification: Generate and purify the wild-type and variant enzymes. Automated systems can be employed for high-throughput processing [83].
- Denaturant Gradient Setup: Using automated liquid handlers, prepare a 24-point gradient of a denaturant such as guanidinium chloride (GdmCl) in a microplate. The concentration range should span from native to fully denaturing conditions.
- Equilibration: Add a fixed concentration of the purified enzyme to each denaturant concentration and incubate to allow the unfolding equilibrium to be reached.
- Fluorescence Measurement: Measure the intrinsic tryptophan fluorescence of the enzyme at each denaturant concentration. The fluorescence signal shifts as the tryptophan residues become exposed to the solvent upon unfolding.
- Data Analysis: Fit the resulting sigmoidal unfolding curve to determine the denaturation midpoint (Cₘ) and the m-value (cooperativity parameter). The stability is then calculated using the linear extrapolation method: ΔG(H₂O) = m * Cₘ [83].
- Comparative Analysis: For mutant enzymes, the change in stability is calculated as ΔΔG = m̄ * (Cₘ(mutant) - Cₘ(WT)), where m̄ is the average m-value of the wild-type and mutant. This method is noted for its precision in high-throughput settings [83].
Live Bacterial Cell Assay for Functional Stability (LEICA)
This innovative, high-throughput method assesses the functional activity and stability of human enzyme variants in an in vivo context using a engineered E. coli chassis, a approach that can be adapted for microbial enzymes [86].
Workflow Overview:
Key Steps:
- Strain Engineering: Create a knockout E. coli strain that lacks a gene essential for metabolism under the assay conditions (e.g., deleted for the gene encoding glucose-6-phosphate isomerase, pgi).
- Genetic Complementation: Transform the knockout strain with plasmids expressing the wild-type or mutant versions of the target enzyme.
- Growth and Selection: Culture the transformed strains in a selective medium (e.g., minimal medium with glucose as the sole carbon source) where robust growth is strictly dependent on the functional activity of the complemented enzyme.
- Growth Kinetics Measurement: Monitor the growth rates of the cultures in a high-throughput manner using automated plate readers.
- Data Correlation: Correlate the measured growth rates with the known activity or stability of the enzyme variants. Studies have demonstrated a high linear correlation (e.g., R² > 0.8) between bacterial growth rate and the biochemically determined activity of recombinant mutant enzymes, validating the assay's accuracy [86].
Computational Methods for Stability Prediction
Computational approaches offer a high-throughput, resource-efficient alternative for predicting the effects of mutations on enzyme stability.
Table 2: Comparison of Computational Methods for Stability Prediction
| Method | Typical Output | Throughput | Key Advantage(s) | Key Limitation(s) |
|---|---|---|---|---|
| Free Energy Perturbation (FEP) | ΔΔG | Low to Medium | Physics-based; high accuracy for relative free energies. | Computationally intensive; requires significant expertise [84]. |
| Machine Learning (AI-based) | Predicted ΔΔG or stability class | Very High | Extremely fast; can leverage large datasets. | Generalizability can be limited; performance depends on training data [84]. |
| Empirical Force Fields (e.g., FoldX) | Predicted ΔΔG | High | Fast; user-friendly. | Lower accuracy compared to more rigorous methods; sensitive to input structure [84]. |
Protocol: Free Energy Perturbation with Hybrid Topology
Modern FEP protocols, such as QresFEP-2, provide a robust physics-based method for calculating the change in thermodynamic stability (ΔΔG) resulting from point mutations [84].
Workflow Overview:
Key Steps:
- Structure Preparation: Obtain a high-resolution crystal or predicted structure of the wild-type enzyme.
- Hybrid Topology Construction: The QresFEP-2 protocol employs a hybrid-topology approach. This involves creating a system where the protein backbone is modeled with a single topology, while the wild-type and mutant side chains are represented by separate, dual topologies. This avoids transforming atom types or bonded parameters, enhancing robustness [84].
- Alchemical Transformation: A coupling parameter (λ) is used to gradually transform the wild-type side chain into the mutant side chain over a series of discrete simulation windows. This is an "alchemical" process not occurring in nature.
- Molecular Dynamics Sampling: Run extensive molecular dynamics simulations at each λ window to adequately sample the conformational space.
- Free Energy Calculation: The free energy change (ΔΔG) is calculated by integrating over the transformation pathway using methods like thermodynamic integration or the Bennett acceptance ratio. The hybrid-topology method is designed for excellent accuracy and computational efficiency, making it suitable for screening hundreds of mutations [84].
The Scientist's Toolkit: Essential Research Reagents and Solutions
Successful stability assessment relies on a suite of specialized reagents and tools.
Table 3: Key Research Reagent Solutions for Stability Assays
| Reagent / Solution | Critical Function | Example Application |
|---|---|---|
| Chemical Denaturants | Unfold protein structure to probe thermodynamic stability. | Guanidinium chloride (GdmCl) or Urea in denaturation curves [83]. |
| Fluorescent Dyes | Report on protein unfolding or aggregation. | Tryptophan fluorescence for intrinsic signal; SYPRO Orange for extrinsic dye-based melting assays [83]. |
| Engineered Microbial Strains | Provide a cellular context for functional stability screening. | E. coli knockout strains (e.g., Δpgi) for the LEICA functional assay [86]. |
| Stabilizing Buffers | Maintain enzyme integrity during purification and assay. | Buffers optimized for extremozymes (e.g., high salt for halophiles) [63] [85]. |
| Immobilization Matrices | Enhance operational stability for repeated-use assays. | Cross-linked enzyme aggregates (CLEAs) for improved thermostability in bioreactors [39]. |
The choice of methodology for comparative enzyme stability assessment is dictated by the project's stage and goals. High-throughput experimental screens like plate-based assays or LEICA are invaluable for initial variant screening and directed evolution campaigns. For detailed mechanistic insights and understanding the thermodynamic basis of stability, lower-throughput methods like detailed denaturation studies are essential. Computational approaches, particularly advanced FEP protocols, are powerful for in silico screening and rational design, bridging the gap between sequence and stability. A robust comparative strategy often integrates multiple methodologies, leveraging the throughput of computation and functional assays with the quantitative precision of rigorous biophysical experiments to deliver a comprehensive stability profile for microbial and extremophile enzymes.
The intrinsic stability of an enzyme at high temperatures is a key determinant of its industrial and research utility. In comparative studies, thermophilic enzymes consistently demonstrate superior thermal stability and a longer functional half-life under thermal stress compared to their mesophilic counterparts. This difference is not merely a function of origin but is rooted in distinct, evolutionarily adapted structural features. Thermophilic enzymes possess a more rigid protein architecture, achieved through a strengthened network of covalent and non-covalent interactions that include an increased number of salt bridges, disulfide bonds, hydrophobic interactions, and hydrogen bonds [87]. Furthermore, their structural composition often features a higher frequency of α-helices and β-sheets in secondary structures and a higher content of certain amino acids like alanine and arginine, all of which contribute to a compact, stable tertiary and quaternary structure that can withstand the atomic vibrations induced by high heat [87] [14].
Conversely, mesophilic enzymes, while often exhibiting higher catalytic activity at moderate temperatures, lack this extensive stabilizing network. Their structures are generally more flexible, which facilitates catalysis at lower temperatures but comes at the cost of being more susceptible to unfolding and irreversible denaturation when the temperature rises [87]. This fundamental trade-off between stability and flexibility is a central theme in understanding the performance differences between these two enzyme classes.
Head-to-Head Comparison: Stability and Kinetic Data
The following tables synthesize experimental data from direct comparative studies, highlighting the distinct performance profiles of thermophilic and mesophilic enzymes.
Table 1: Comparative Thermal Stability and Half-Life of Model Enzymes
| Enzyme (Organism) | Enzyme Class | Thermal Stability (Melting Temperature, Tₘ) | Half-Life (t₁/₂) | Experimental Conditions |
|---|---|---|---|---|
| 3-Phosphoglycerate Kinase (Thermoanaerobacter sp.) | Thermophilic | Not Specified | Apparent Eact for denaturation: 600 kJ·mol⁻¹ [88] | Dilute buffer |
| 3-Phosphoglycerate Kinase (Zymomonas mobilis) | Mesophilic | Not Specified | Apparent Eact for denaturation: 200-250 kJ·mol⁻¹ [88] | Dilute buffer |
| WF146 Protease (Thermophilic Bacillus sp.) | Thermophilic (Subtilase) | Tₘ of variant PBL5X: 5.5°C higher than wild-type [89] | Wild-type: 6.3 min at 85°CVariant PBL5X: 57.1 min at 85°C [89] | Purified enzyme in buffer |
| Subtilisin S41 (Psychrophilic) | Psychrophilic (Subtilase) | Low | Expected to be very short at high temperatures | Purified enzyme |
Table 2: Comparative Kinetic Parameters at Different Temperatures
| Enzyme (Organism) | Optimal Temperature (Tₒₚₜ) | kcat at Lower Temperatures | Catalytic Efficiency | Key Kinetic Observation |
|---|---|---|---|---|
| 3-Phosphoglycerate Kinase (Thermophile) | Close to organism's optimum (~68°C) [88] | Lower than mesophiles [88] | Maintains efficiency at high T | Km values stable at lower T, increase substantially after Tₒₚₜ [88] |
| 3-Phosphoglycerate Kinase (Mesophile) | Much higher than organism's optimum (~32°C) [88] | Higher than thermophiles at low T [88] | Loses efficiency at high T | Km values stable at lower T, increase substantially after Tₒₚₜ [88] |
| TtIPMDH (Thermus thermophilus) | High | Low at 25°C [90] | Lower at low T | Inverse correlation between stability and low-T activity [90] |
| EcIPMDH (Escherichia coli) | Moderate | High at 25°C [90] | Higher at low T | Inverse correlation between stability and low-T activity [90] |
| TtIPMDH Mutant 9/21 (Engineered) | Retained high stability [90] | 17x higher than wild-type Tt at 25°C [90] | Significantly improved | Demonstrated breaking of stability-activity trade-off via engineering [90] |
Experimental Protocols for Key Findings
The data presented in the comparison tables are derived from rigorous experimental methodologies. Below are the protocols for two critical experiments that underscore the differences between thermophilic and mesophilic enzymes.
Protocol: Measuring Irreversible Thermal Denaturation
This method was used to generate the half-life and activation energy (Eact) data for phosphoglycerate kinase in Table 1 [88].
- Enzyme Purification: Purify the target enzyme (e.g., 3-phosphoglycerate kinase) from both thermophilic and mesophilic organisms using standard chromatographic techniques.
- Activity Assay Setup: Establish a standardized continuous assay to monitor the catalytic activity of the enzyme. For kinases, this typically involves coupling the reaction to the oxidation of NADH and measuring the decrease in absorbance at 340 nm.
- Thermal Denaturation: Incubate separate aliquots of the purified enzyme in a dilute buffer at a constant, elevated temperature in a thermal block.
- Sampling: At regular time intervals, remove samples from the heating block and immediately place them on ice to halt denaturation.
- Residual Activity Measurement: Assay the remaining catalytic activity of the cooled samples using the standardized assay from step 2.
- Data Analysis: Plot the logarithm of the residual activity against the incubation time. The half-life (t₁/₂) is determined as the time at which 50% of the initial activity is lost. The apparent activation energy for denaturation (Eact) can be calculated from an Arrhenius plot of the denaturation rate constants obtained at different temperatures.
Protocol: Engineering a Thermophilic Enzyme for Enhanced Low-Temperature Activity
This rational design approach, which produced the TtIPMDH mutant data in Table 2, involves systematically introducing mesophilic features into a thermophilic enzyme [90].
- Target Identification:
- Perform a pairwise sequence alignment of a thermophilic enzyme (e.g., TtIPMDH from Thermus thermophilus) with its mesophilic homologue (e.g., EcIPMDH from E. coli).
- Identify non-conserved amino acid residues located within a defined radius (e.g., 8-12 Å) of the enzyme's active site.
- Site-Directed Mutagenesis:
- Create a series of mutant constructs where single or groups of target residues in the thermophilic enzyme are replaced with the residues found at the equivalent positions in the mesophilic enzyme.
- This can be achieved using techniques like the QuikChange method or overlap extension PCR.
- Expression and Purification:
- Heterologously express the wild-type and mutant enzymes in a suitable host (e.g., E. coli).
- Purify the proteins to homogeneity using affinity chromatography to ensure comparable samples.
- High-Throughput Screening:
- Measure the specific activity of all mutant enzymes at the target low temperature (e.g., 25°C).
- Identify "hits" – mutants that show significantly improved activity over the wild-type thermophilic enzyme.
- Combination and Characterization:
- Combine beneficial mutations from different hits into a single construct.
- Thoroughly characterize the kinetic parameters (kcat, Km) and thermal stability (Tₘ, t₁/₂) of the best-performing combined mutant to confirm enhanced low-temperature activity without a significant loss of thermostability.
Stability and Catalytic Mechanism Workflows
The relationship between an enzyme's structural stability and its catalytic function is complex. The following diagrams illustrate the conceptual trade-off and a modern method to probe it experimentally.
The Activity-Stability Trade-off
Enzyme Proximity Sequencing (EP-Seq) Workflow
The Scientist's Toolkit: Key Research Reagents and Solutions
Table 3: Essential Reagents for Enzyme Stability and Activity Research
| Reagent / Solution | Function / Application | Example in Context |
|---|---|---|
| Affinity Chromatography Resins | Purification of recombinant enzymes, often via His-tags or substrate analogues. | Bacitracin-Sepharose for subtilase purification [89]; Ni²⁺-charged resin for His-tagged proforms [89]. |
| Stabilizing Buffers and Cofactors | Maintaining enzyme stability during purification and assays; essential for activity. | Buffers with CaCl₂ for subtilases [89]; NAD⁺ for dehydrogenases [90]; FAD for oxidases like DAOx [29]. |
| Chemical Substrates and Analogs | Measuring enzyme kinetics and probing active site structure. | suc-AAPF-pNA for subtilase activity [89]; 3-IPM for IPMDH kinetics [90]; CdRP for IGPS studies [91]. |
| Protease Inhibitors (e.g., PMSF) | Halting unwanted proteolytic degradation during enzyme purification and processing. | Used to inactivate residual subtilisin A after processing proforms of WF146 protease [89]. |
| Fluorescent Conjugates for Assays | Enabling high-throughput screening and deep mutational scanning. | Anti-His antibodies for expression staining; Tyramide-488 for proximity labeling in EP-Seq [29]. |
| Specialized Expression Systems | Heterologous production of extremozymes, which can be challenging in standard hosts. | E. coli BL21(DE3) for recombinant protein expression [89]; Yeast surface display for EP-Seq [29]. |
Tolerance to Organic Solvents, Extreme pH, and High Salt Concentrations
Enzymes are indispensable biocatalysts in numerous industrial and pharmaceutical processes. However, their widespread application is often constrained by a lack of stability under harsh operational conditions, including the presence of organic solvents, extreme pH, and high salt concentrations [92] [93]. While enzymes from mesophilic microorganisms (microbial enzymes) are commonly used, they frequently denature and lose activity in such non-conventional environments [94]. In contrast, enzymes derived from extremophilic microorganisms (extremozymes) have evolved unique structural adaptations that confer remarkable resilience, making them superior candidates for demanding applications [95] [2]. This guide provides an objective comparison of the stability and performance of microbial versus extremophile enzymes, presenting key experimental data to inform researchers and drug development professionals.
Comparative Enzyme Stability: Quantitative Data
The following tables summarize experimental data comparing the stability of various microbial and extremophile enzymes under different stress conditions.
Table 1: Comparative Stability of Enzyme Classes Under Stress Conditions
| Enzyme Class | Source Organism | Organic Solvent Tolerance (e.g., Log P value) | pH Stability Range (Optimum) | Salt Tolerance (e.g., NaCl Concentration) | Half-life/Activity Retention |
|---|---|---|---|---|---|
| Psychrophilic Protease | Antarctic Bacteria [3] | Moderate | 5.0-9.0 [2] | Moderate | High activity at 5°C [3] |
| Thermophilic Protease | Bacillus subtilis [2] | Moderate | 9.0 (Optimum) [2] | High (Halotolerant) [2] | ~4 hours at 60°C [96] |
| Thermophilic α-Amylase | Thermus species [95] | High | 5.0-8.0 [93] | Low-Moderate | Stable at 50-80°C [93] |
| Halophilic α-Amylase | Halotolerant Bacteria [2] | High | 6.0-10.0 [2] | Very High (≥4 M NaCl) [2] | Enhanced activity at high salt [2] |
| Mesophilic Lipase | Pseudomonas sp. [92] | Low-Moderate (Log P >4 preferred) [92] | Narrow (Varies) | Low | Rapid inactivation under extremes [92] |
| Solvent-Tolerant Lipase | Aneurinibacillus sp. [92] | Very High (Tolerant to >30% solvents) [92] | Broad (Varies) | Moderate-High | Stable in hydrophobic solvents [92] |
| Phosphite Dehydrogenase (PtxD) | Ralstonia sp. 4506 (Soil Bacterium) [94] | Low (Inhibited by 10-30% solvents) [94] | - | Low | Susceptible to salt ions [94] |
| Phosphite Dehydrogenase (PtxD) | Cyanothece sp. ATCC 51142 (Marine Cyanobacterium) [94] | Very High (Enhanced activity in 10-30% solvents) [94] | 6.0-10.0 (Optimum at 9.0) [94] | Very High (Resistant to Na+, K+, NH4+) [94] | Stable over broad pH range; optimal at 50°C [94] |
Table 2: Experimental Performance Data in Biocatalytic Reactions
| Enzyme | Reaction Condition Stressors | Performance Metric (e.g., Conversion Rate, Yield) | Comparison to Microbial Counterpart |
|---|---|---|---|
| Ct-PtxD (Extremophile) | High Ammonium Conditions [94] | Efficient NADH regeneration for L-tert-leucine synthesis [94] | Superior to Rs-PtxD, which is less supported [94] |
| Solvent-Stable Lipase | Non-aqueous media (e.g., synthesis of esters) [92] | High catalytic efficiency, shifts equilibrium to synthesis [92] | Outperforms mesophilic lipases, which denature [92] |
| Halophilic Protease | High Salt, Alkaline pH (Detergent conditions) [2] | Maintains high activity for stain removal [2] | More stable than standard bacterial proteases [2] |
The superior stability of extremozymes is not random but stems from specific evolutionary adaptations at the molecular level.
Organic Solvent Tolerance: Solvents, particularly polar ones (log P < 2), strip essential water molecules from enzymes, leading to denaturation [92]. Extremozymes combat this through strategies such as a more rigid and compact structure, a higher surface charge density to maintain a hydration shell, and a reduced surface hydrophobic amino acid content to minimize unfavorable solvent interactions [92] [94]. For instance, the solvent-tolerant phosphite dehydrogenase (Ct-PtxD) from Cyanothece has fewer hydrophobic surface residues, reducing aggregation and denaturation in low-water-activity environments [94].
Extreme pH Tolerance: Activity at non-neutral pH is enabled by an altered surface charge distribution. Acidophilic enzymes have a high density of acidic residues, while alkaliphilic enzymes possess more basic residues, ensuring stability and correct ionization of the active site across the pH spectrum [2]. This prevents irreversible conformational changes and loss of activity.
High Salt Concentration Tolerance: High ionic strength can screen electrostatic interactions critical for protein folding and function. Halophilic enzymes adapt by having a high content of acidic residues (aspartate and glutamate) on their surface, which coordinate hydrated ions and maintain a water shell—a process known as "solvation by salt" [2] [94]. This keeps the enzyme hydrated and functional at near-saturation salt conditions.
The following diagram illustrates the core mechanistic relationships that confer stability in extremophile enzymes.
Mechanisms of Extremozyme Stability under Stress Conditions.
Detailed Experimental Protocols
To objectively compare enzyme stability, standardized assays are employed. Below are detailed methodologies for key experiments cited in this guide.
Protocol: Assessing Organic Solvent Tolerance
This protocol is adapted from studies on solvent-stable lipases and Ct-PtxD [92] [94].
- Objective: To determine the effect of organic solvents on enzyme activity and stability.
- Materials:
- Purified enzyme solution.
- Organic solvents of varying log P values (e.g., ethanol, dimethylformamide (DMF), methanol, toluene).
- Standard assay buffers and substrates specific to the enzyme (e.g., p-nitrophenyl ester for lipases).
- Thermostatic incubator.
- Spectrophotometer or HPLC for product quantification.
- Method:
- Preparation of Solvent-Enzyme Mixtures: In glass vials, mix the enzyme solution with the organic solvent to achieve final solvent concentrations (e.g., 10%, 20%, 30% v/v). Use a vial with no solvent as a control.
- Incubation: Incubate the mixtures at a constant temperature (e.g., 30°C or 50°C) with gentle shaking for a predetermined period (e.g., 1, 6, 24 hours).
- Residual Activity Assay: After incubation, remove an aliquot from each vial. For water-miscible solvents, the aliquot can be diluted directly into the standard activity assay mixture. For water-immiscible solvents, use the aqueous phase. Measure the initial rate of the enzymatic reaction.
- Calculation: Calculate the residual activity as a percentage of the activity of the unincubated control (or the control incubated without solvent). Plot residual activity vs. solvent type/concentration.
Protocol: Assessing pH and Salt Stability
This protocol is based on characterizations of halophilic and alkaliphilic enzymes [2] [94].
- Objective: To determine the optimal pH and salt concentration for enzyme activity and its stability over a broad range.
- Materials:
- Purified enzyme.
- A series of buffers covering a broad pH range (e.g., pH 3-11, such as citrate, phosphate, Tris, glycine).
- Salt solutions (e.g., NaCl, KCl, (NH₄)₂SO₄) at various molarities.
- Standard assay reagents.
- Method:
- Optimal pH: Prepare the standard activity assay mixture using buffers of different pH values. Initiate the reaction by adding the enzyme and measure the initial reaction rate. Plot activity vs. pH to determine the pH optimum.
- pH Stability: Incubate the enzyme in different pH buffers without substrate for a fixed time (e.g., 1 hour). Then, adjust the pH to the optimum condition or dilute into the standard assay (which is at the optimal pH) and measure the residual activity.
- Salt Dependence and Tolerance: Perform the standard activity assay in the presence of different concentrations of salt. Plot activity vs. salt concentration. To test salt stability, pre-incubate the enzyme with high salt concentrations before assaying under standard conditions.
The Scientist's Toolkit: Key Research Reagents and Solutions
Table 3: Essential Reagents for Studying Enzyme Stability
| Reagent / Material | Function / Application | Examples / Notes |
|---|---|---|
| Extremophile Cell Lines | Source of extremozymes for discovery and purification. | Halotolerant Bacillus subtilis CH11 [96], Marine Cyanothece sp. [94], Thermophilic Pyrococcus furiosus [93]. |
| Heterologous Expression Hosts | Production of recombinant extremozymes. | Escherichia coli Rosetta2 (DE3) for Ct-PtxD expression [94]. |
| Organic Solvents (Varying Log P) | To create low water activity environments and test solvent tolerance. | Polar (e.g., Methanol, DMF, Ethanol) and Non-polar (e.g., Toluene, Hexane) [92]. |
| Salt Ions | To create high ionic strength conditions and test halotolerance. | NaCl, KCl, NH₄⁺ [94]. Specific ions can have varying effects. |
| Immobilization Carriers | Enhancing enzyme stability, reusability, and performance under harsh conditions. | Chitosan, alginate [11]; Bacterial spores (Bacillus subtilis) as safe, effective display system [59]. |
| Cross-linking Reagents | For enzyme immobilization via covalent bonding. | Glutaraldehyde, carbodiimide [11]. |
The experimental data and comparative analysis presented in this guide objectively demonstrate the superior robustness of extremophile enzymes under conditions of organic solvents, extreme pH, and high salt concentrations compared to their conventional microbial counterparts. While microbial enzymes are effective under mild conditions, their instability in harsh industrial processes limits their utility. The innate structural and functional adaptations of extremozymes not only provide a natural solution to these challenges but also reduce the need for extensive process engineering or protein modification. For researchers and drug development professionals aiming to develop efficient and sustainable biocatalytic processes, the targeted selection or discovery of extremozymes is a strategically advantageous path forward.
Comparative Analysis of Operational Stability in Simulated Industrial Processes
Operational stability is a critical determinant of efficiency and cost-effectiveness in industrial biotechnology, particularly in processes involving enzymatic reactions. This guide provides a comparative analysis of the operational stability between traditional microbial enzymes and extremophile-derived enzymes (extremozymes) within simulated industrial environments. The performance of these biocatalysts is evaluated based on their resilience to extreme operational parameters such as temperature, pH, and organic solvents—conditions often encountered in manufacturing pharmaceuticals, fine chemicals, and other high-value products. For researchers and drug development professionals, selecting the appropriate enzyme type is paramount for process robustness, reduced downtime, and overall economic viability. This analysis synthesizes experimental data and mechanistic insights to offer an evidence-based comparison, framing the discussion within the broader context of advancing biocatalytic stability for industrial applications.
Performance Comparison: Microbial Enzymes vs. Extremozymes
The following tables summarize key stability parameters and industrial performance metrics for microbial and extremophile enzymes, based on experimental findings and biotechnological applications.
Table 1: Comparative Structural and Functional Stability Parameters
| Stability Parameter | Microbial (Mesophilic) Enzymes | Extremophile-Derived Enzymes (Extremozymes) | Experimental Conditions & Notes |
|---|---|---|---|
| Thermal Stability (Half-life) | Typically minutes to a few hours at >60°C [2] | Several hours to days at 60-100°C+ [2] [50] | Measured via incubation at 70°C; thermophiles show enhanced salt bridges & H-bonds [2]. |
| Thermal Inactivation Point | Often denatures above 50-60°C [4] | Retains function at 80-122°C (thermophiles) [2] [4] | Optimal Growth Temperature (OGT) correlates with amino acid composition bias [2]. |
| Low-Temperature Activity | Activity significantly decreases [2] | High catalytic efficiency at 0-15°C (psychrophiles) [2] [4] | Psychrophilic enzymes have low-activation enthalpy and heat-labile active sites [2]. |
| pH Stability Range | Generally narrow, optimal near pH 7 | Stable and active at pH <4 (acidophiles) or >9 (alkaliphiles) [50] [4] | Alkaliphiles use cytochrome c and cell wall proteins to manage proton gradient [4]. |
| Solvent/Oxidant Tolerance | Moderate; often requires process optimization | High intrinsic tolerance to organic solvents & oxidants [50] | Polyextremophilic features enable function in harsh industrial conditions [50]. |
Table 2: Industrial Performance and Economic Metrics
| Performance Metric | Microbial (Mesophilic) Enzymes | Extremophile-Derived Enzymes (Extremozymes) | Relevant Industrial Applications |
|---|---|---|---|
| Operational Half-life | Shorter; necessitates frequent replacement or replenishment [2] | Prolonged; significantly reduces enzyme consumption and cost-in-use [50] | Continuous biotransformation processes in pharmaceutical synthesis. |
| Process Cost Efficiency | Lower for standard conditions; high for extreme conditions due to supporting infrastructure | Higher for extreme conditions; reduces need for cooling, heating, or pH adjustment [50] | Starch processing (thermostable amylases), detergents (alkaliphilic proteases) [2] [50]. |
| Catalytic Efficiency (kcat/Km) | Optimized for mild, physiological conditions | Can be significantly higher under non-standard conditions (e.g., cold-active enzymes at 10°C) [2] | Food processing (psychrophilic enzymes for low-temperature fermentation) [2]. |
| Reaction Output Stability | Can fluctuate with minor process deviations | Highly consistent output across varied and fluctuating bioreactor conditions [50] | High-fructose syrup production using thermostable invertase [50]. |
Experimental Protocols for Assessing Operational Stability
To generate comparable data on enzyme stability, standardized experimental protocols are essential. The following methodologies are widely used for evaluating key performance parameters.
Protocol for Thermal Stability and Half-life Determination
Objective: To quantify an enzyme's resistance to thermal denaturation over time and determine its half-life at a specified temperature.
- Sample Preparation: Prepare a purified enzyme solution at a standardized concentration in an appropriate buffer. Aliquot the solution into multiple, identical PCR tubes.
- Incubation: Place the aliquots in a thermocycler or precision water bath pre-set to the target temperature (e.g., 60°C, 70°C, 80°C). Include a control sample kept on ice (0°C).
- Sampling: At predetermined time intervals (e.g., 0, 5, 15, 30, 60, 120 minutes), remove one aliquot from the heating block and immediately transfer it to ice to halt thermal inactivation.
- Residual Activity Assay: For each time-point sample, measure the remaining enzymatic activity using a standardized assay under optimal conditions (e.g., 37°C for mesophiles). The activity of the time-zero sample represents 100% initial activity.
- Data Analysis: Plot the natural logarithm of residual activity (%) versus time. The half-life (t1/2) is calculated from the slope of the linear regression (k, the inactivation rate constant) using the formula: t1/2 = ln(2) / k. A longer half-life indicates superior thermal stability [2].
Protocol for pH Stability Profiling
Objective: To determine the range of pH over which an enzyme retains its structural integrity and functional activity.
- Buffer System Setup: Prepare a series of buffers covering a broad pH range (e.g., pH 2-12). Use appropriate buffer substances with overlapping pKa values to ensure consistent buffering capacity.
- Enzyme Exposure: Incubate separate aliquots of the purified enzyme in each buffer solution for a fixed duration (e.g., 1 hour) at a constant, non-denaturing temperature (e.g., 25°C).
- Activity Measurement: After incubation, adjust the pH of each aliquot to the enzyme's known pH optimum or perform the activity assay in a universal buffer at the optimum pH. This step ensures that the measured activity reflects pH-induced stability loss, not just transient pH effects on catalysis.
- Analysis: Express the activity at each pre-incubation pH as a percentage of the maximum activity observed. The resulting profile shows the pH range for operational stability, with extremozymes typically displaying broader or shifted profiles compared to mesophilic enzymes [50] [4].
Protocol for Solvent Tolerance Assessment
Objective: To evaluate enzyme activity and stability in the presence of water-miscible organic solvents, which is critical for non-aqueous biocatalysis.
- Reaction Mixture Preparation: Set up reaction mixtures containing the enzyme, its substrate, and a buffer with varying concentrations (e.g., 10%, 20%, 30% v/v) of an organic solvent like methanol, dimethyl sulfoxide (DMSO), or acetonitrile. A control reaction with no solvent is included.
- Initial Rate Measurement: Immediately measure the initial reaction rate for each mixture. This indicates the solvent's effect on catalytic activity.
- Long-term Stability Incubation: In a separate experiment, incubate the enzyme in buffer-solvent mixtures (without substrate) for several hours.
- Residual Activity Measurement: After incubation, dilute the samples to minimize the solvent concentration and assay for residual activity relative to a control incubated without solvent. This measures the solvent's effect on long-term structural stability. Enzymes from halophiles or thermophiles often show superior retention of activity [50].
Mechanisms of Stability: A Comparative Workflow
The fundamental difference in operational stability originates from distinct evolutionary adaptations encoded in the enzyme's structure. The following diagram and explanation outline the key mechanistic pathways.
Mechanistic Workflow of Enzyme Stability:
The workflow is initiated by Environmental Stress, such as high temperature, extreme pH, or high salinity. In response, Molecular Adaptation Mechanisms are engaged. These mechanisms differ profoundly between standard microbial enzymes and extremozymes.
Microbial (Mesophilic) Enzymes typically possess only limited adaptations, resulting in outcomes such as a narrow operational range and lower structural stability under extreme conditions. Their structures are optimized for the mild, stable environments of their host organisms.
Extremozymes exhibit specialized adaptations refined through evolution in extreme niches. For thermophiles, this involves a higher number of salt bridges, hydrogen bonds, hydrophobic core interactions, and more compact protein structures with shortened loops to resist denaturation [2]. For psychrophiles, adaptations include greater molecular flexibility, a reduced hydrophobic core, and weaker inter-domain interactions to maintain activity at low temperatures [2] [4]. Acidophiles and alkaliphiles feature specialized membrane transporters and surface protein networks to maintain internal pH homeostasis [4]. These specialized mechanisms lead to the outcome of a wide operational range and high stability under conditions that would inactivate mesophilic enzymes.
The Scientist's Toolkit: Essential Reagents and Solutions
Table 3: Key Reagents for Enzyme Stability Research
| Reagent / Material | Function in Experimental Protocol | Specific Application Example |
|---|---|---|
| Recombinant Extremozymes | The core biocatalyst for testing; produced via heterologous expression in mesophilic hosts (e.g., E. coli, Komagataella pastoris) [50]. | Thermostable invertase from Gongronella sp. for high-fructose syrup production [50]. |
| Broad-Range Buffer Systems | To create and maintain specific pH environments during stability profiling. | Assessing protease activity across pH 4-11 to determine suitability for detergent formulations [50]. |
| Compatible Solutes (e.g., trehalose, betaine) | To stabilize enzyme structure against denaturing stresses like heat, freezing, or desiccation [4]. | Added to lyophilization buffers to enhance the shelf-life of enzyme formulations. |
| Immobilization Resins (e.g., epoxy, chitosan) | To covalently or physically bind enzymes onto solid supports, enhancing their operational stability and enabling reusability. | Cellulose-based resins for immobilizing invertase in continuous-flow bioreactors [50]. |
| Organic Solvents (HPLC grade) | To create water-miscible or biphasic systems for solvent tolerance assays. | Evaluating lipase activity in non-aqueous synthesis of chiral pharmaceutical intermediates. |
| Spectrophotometric Assay Kits | To quantitatively measure enzyme activity via the detection of a colored or fluorescent product. | Monitoring the hydrolysis of a p-nitroanilide-linked substrate by a protease. |
The comparative analysis unequivocally demonstrates that extremozymes offer superior operational stability under a wide spectrum of extreme industrial conditions compared to their mesophilic counterparts. While traditional microbial enzymes are effective for processes operating within mild parameters, the intrinsic structural robustness of extremozymes—derived from specific molecular adaptations—translates into longer operational half-lives, greater process consistency, and reduced operational costs in demanding applications. For researchers and drug development professionals, the strategic integration of extremozymes into process design represents a forward-looking approach to building more resilient, efficient, and sustainable industrial bioprocesses. The continued exploration of extremophilic biodiversity and advancement in protein engineering will further expand the toolbox of stable biocatalysts, pushing the boundaries of industrial biotechnology.
The escalating demand for sustainable industrial processes has catalyzed a shift from traditional chemical catalysts to biological alternatives, with enzymes emerging as pivotal tools. However, conventional enzymes derived from mesophilic organisms often exhibit limited stability under the harsh conditions typical of industrial manufacturing, such as extreme temperatures, pH, and the presence of organic solvents. This instability can lead to frequent enzyme replacement, high operational costs, and significant process inefficiencies. Extremozymes, derived from microorganisms that thrive in extreme environments, present a transformative solution to these challenges. These enzymes are characterized by their innate robustness, maintaining structural integrity and catalytic efficiency under conditions that would denature their mesophilic counterparts. This analysis provides a comparative evaluation of the economic and environmental impacts of adopting extremozymes, benchmarking their performance against traditional microbial enzymes to outline a compelling case for their integration into modern industrial workflows.
Performance and Stability Comparison: Extremozymes vs. Microbial Enzymes
The operational advantages of extremozymes are most evident in their performance under extreme physicochemical conditions. The data below quantitatively compares key stability parameters between extremozymes and conventional microbial enzymes.
Table 1: Comparative Stability of Extremozymes vs. Conventional Microbial Enzymes
| Parameter | Extremozymes | Conventional Microbial Enzymes | Industrial & Environmental Impact |
|---|---|---|---|
| Thermal Stability | Stable at temperatures >100°C (e.g., Methanopyrus kandleri at 122°C) [18] | Typically denature above 40-70°C | Enables higher temperature processes, reduces cooling costs, lowers risk of microbial contamination [18] [97] |
| pH Tolerance | Functional in highly acidic (pH 0-2) or alkaline (pH 9-12) ranges [18] | Optimal activity in near-neutral pH ranges (pH 5-8) | Eliminates need for pH adjustment steps, reduces chemical consumption and effluent salinity [18] |
| Organic Solvent Stability | Retain function in non-aqueous and organic solvent systems [98] | Often precipitate or denature | Facilitates synthesis of chiral pharmaceuticals and fine chemicals in organic media [98] |
| Operational Half-Life | Significantly extended due to inherent robustness; further enhanced via immobilization [99] | Relatively short, requires frequent replenishment | Reduces enzyme consumption, lowers raw material costs, and minimizes process downtime [100] [99] |
| Catalytic Efficiency | High specific activity at low temperatures (psychrophiles) or high temperatures (thermophiles) [97] | Activity drops sharply outside narrow optimal ranges | Increases reaction velocity, improves product yield, and allows for smaller reactor volumes [97] |
Experimental Analysis of Comparative Stability
Experimental Protocol for Assessing Thermostability
Objective: To quantitatively compare the thermal stability of a hyperthermophilic protease (e.g., from Pyrococcus furiosus) with a mesophilic protease (e.g., from Bacillus licheniformis).
Methodology:
- Enzyme Preparation: Prepare purified solutions of both enzymes in appropriate buffers at a standardized concentration (e.g., 1 mg/mL).
- Heat Incubation: Aliquot enzyme solutions and incubate them at a elevated temperature (e.g., 70°C, 80°C, 90°C). Remove samples at defined time intervals (e.g., 0, 15, 30, 60, 120 minutes).
- Residual Activity Assay: Immediately cool samples on ice. Measure residual enzymatic activity using a standard protease assay (e.g., casein hydrolysis at 37°C or 60°C). The reaction mixture typically contains substrate (casein), buffer, and the quenched enzyme sample. The reaction is stopped by adding trichloroacetic acid, and the solubilized tyrosine is measured spectrophotometrically at 280 nm.
- Data Analysis: Express residual activity as a percentage of the initial activity (t=0 sample). Plot residual activity versus incubation time to determine the half-life of each enzyme at the various temperatures.
Expected Outcome: The hyperthermophilic protease will exhibit a significantly longer half-life at high temperatures, with minimal activity loss over several hours, whereas the mesophilic protease will rapidly denature, losing most of its activity within minutes [18] [97].
Experimental Protocol for Assessing pH Stability
Objective: To evaluate the stability and activity of an acidophilic enzyme (e.g., a xylanase from Saccharolobus solfataricus) across a broad pH range compared to a commercial fungal xylanase.
Methodology:
- pH Profiling: Prepare a series of buffers covering a wide pH range (e.g., pH 2.0 to 10.0).
- Pre-incubation Stability: Incubate each enzyme in the different pH buffers for a fixed period (e.g., 1 hour) at a moderate temperature (e.g., 25°C). Then, adjust the pH to the optimal pH for the activity assay to measure residual activity.
- Activity vs. pH: Directly assay enzyme activity at each pH without a pre-incubation step to determine the pH-activity profile. The activity assay can use birchwood xylan as a substrate, with reducing sugars measured by the DNS method.
- Data Analysis: Plot both residual activity (stability) and initial activity versus pH.
Expected Outcome: The acidophilic xylanase will maintain high stability and activity in the acidic pH range (e.g., pH 2-4), while the commercial fungal enzyme will show a narrow pH optimum and rapid inactivation at acidic pH [18] [4].
Economic Cost-Benefit Analysis
The initial higher cost of extremozyme production is offset by substantial long-term savings and environmental benefits, creating a compelling economic case.
Table 2: Comprehensive Cost-Benefit Analysis of Extremozyme Adoption
| Factor | Extremozymes | Conventional Microbial Enzymes | Remarks |
|---|---|---|---|
| Upfront R&D & Production Cost | High (specialized fermentation, downstream processing) [48] | Low (established fermentation protocols) | AI-driven enzyme design is reducing development timelines from 3-5 years to 6-18 months, cutting costs by 50-70% [100]. |
| Operational Cost | Lower (longer half-life, reusability via immobilization) [99] | Higher (frequent replenishment needed) | Immobilization allows for enzyme reuse over multiple cycles, drastically reducing cost per unit of product [100] [99]. |
| Process Efficiency | Higher (faster kinetics, higher temperature operation, less downtime) [97] | Lower | Enables continuous processing and reduced reactor sizing due to faster reaction rates [100]. |
| Energy Consumption | 50-70% reduction compared to chemical processes [100] | Moderate savings | High thermal stability eliminates need for energy-intensive cooling/heating cycles to maintain enzyme function [97]. |
| Environmental Compliance Cost | Lower (biodegradable, reduced toxic waste) [18] [12] | Variable | Replaces harsh chemicals in processes like textile processing (e.g., chlorine bleach, harsh acids/alkalis), simplifying effluent treatment [100] [12]. |
Molecular Basis of Extremozyme Stability
The remarkable stability of extremozymes is not due to a single factor but arises from a suite of sophisticated structural adaptations. The following diagram illustrates the key molecular strategies that confer stability across different types of extremozymes.
The Scientist's Toolkit: Key Research Reagents and Solutions
Advancing research and development in extremozyme technology requires a specific set of specialized reagents and tools.
Table 3: Essential Research Reagents for Extremozyme Investigation
| Reagent / Material | Function & Application | Specific Example |
|---|---|---|
| Specialized Growth Media | Cultivating extremophilic microorganisms under simulated native conditions (e.g., high salt, extreme pH, elevated temperature). | Media with high NaCl (≥15%) for halophiles; low pH (2.0) buffers for acidophiles; anaerobic chambers for anaerobes [18] [98]. |
| Activity Assay Kits | Quantifying enzymatic activity under extreme conditions using chromogenic/fluorogenic substrates. | Protease assay using casein/azocasein at 80°C; cellulase assay using carboxymethyl cellulose at pH 2.0 [98]. |
| Protein Stabilizers | Maintaining enzyme integrity during purification, storage, and handling. | Compatible solutes (e.g., ectoine, betaine), polyols (e.g., glycerol), and salts [98]. |
| Immobilization Matrices | Enhancing enzyme reusability, stability, and simplifying product separation. | Functionalized silica, chitosan, magnetic nanoparticles, and epoxy-activated resins [100] [99]. |
| Cloning & Expression Systems | Genetic manipulation and heterologous production of extremozymes. | Genetically engineered thermophilic hosts (e.g., Thermus thermophilus), E. coli with codon optimization, and specialized vectors [48]. |
The integration of extremozymes into industrial biocatalysis represents a paradigm shift towards a more sustainable and economically competitive bioeconomy. While the initial investment in their discovery and production is non-trivial, the long-term benefits—marked by enhanced process robustness, significant operational savings, and a dramatically reduced environmental footprint—present an undeniable value proposition. The experimental data and cost analysis confirm that the superior intrinsic stability of extremozymes translates directly into tangible economic and environmental advantages. As protein engineering and metagenomic screening technologies continue to advance, the gap between the potential and application of extremozymes will narrow, accelerating their adoption across sectors from pharmaceuticals and biofuels to textile manufacturing and environmental remediation.
Conclusion
The comparative analysis conclusively demonstrates that extremozymes possess superior and often specialized stability under harsh conditions compared to conventional microbial enzymes, making them invaluable for biomedical and industrial applications. Their inherent resilience to temperature, pH, and solvents directly translates to more efficient, sustainable, and cost-effective biocatalytic processes. For drug development, this opens avenues for novel therapeutics and more robust manufacturing pipelines. Future research must focus on overcoming production bottlenecks through advanced synthetic biology and metagenomic exploration. Integrating these robust biocatalysts will be pivotal for advancing green chemistry, developing next-generation pharmaceuticals, and pushing the boundaries of biotechnological innovation.
References
- 1. Enhancing stability of enzymes for industrial applications [https://link.springer.com/article/10.1007/s11274-025-04568-4]
- 2. Extremophile – An Adaptive Strategy for Extreme Conditions ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7324872/]
- 3. Advances in Extremophile Research: Biotechnological ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11433292/]
- 4. Extremophiles and Extremophilic Behaviour—New Insights ... [https://www.mdpi.com/2075-1729/14/11/1425]
- 5. Microbial enzymes and major applications in the food industry [https://fppn.biomedcentral.com/articles/10.1186/s43014-024-00261-5]
- 6. Extremophiles: Unlocking biomedical and industrial ... [https://www.cas.org/resources/cas-insights/extremophiles-biomedical-industrial-innovations]
- 7. Enzyme Miniaturization: Revolutionizing Future Biocatalysts [https://pmc.ncbi.nlm.nih.gov/articles/PMC12226312/]
- 8. Enzyme specificity prediction using cross-attention graph ... [https://www.nature.com/articles/s41586-025-09697-2]
- 9. Microbial Enzymes: Tools for Biotechnological Processes [https://pmc.ncbi.nlm.nih.gov/articles/PMC4030981/]
- 10. Stability and adaptation of proteins, membranes and cells ... [https://www.sciencedirect.com/science/article/abs/pii/S0022283625005431]
- 11. Review of research progress in immobilization and chemical ... [https://microbialcellfactories.biomedcentral.com/articles/10.1186/s12934-025-02791-0]
- 12. Recent Advances in Microbial Enzyme Applications for ... [https://www.mdpi.com/2413-4155/7/2/46]
- 13. Exploring Bacterial Interactions Under the Stress Gradient ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12413402/]
- 14. Extremophile Microbial Communities and Enzymes for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7521039/]
- 15. A Perspective Towards More Sustainable Production of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12525642/]
- 16. The Extremophiles: Adaptation Mechanisms and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12024854/]
- 17. How Extremophiles Push the Limits of Life [https://asm.org/articles/2023/march/how-extremophiles-push-the-limits-of-life]
- 18. Biotechnological Potential of Extremophiles: Environmental ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12292952/]
- 19. Factors enhancing protein thermostability [https://pubmed.ncbi.nlm.nih.gov/10775659/]
- 20. Neural network conditioned to produce thermophilic ... protein [https://www.nature.com/articles/s41598-025-90828-0?error=cookies_not_supported&code=3d4d9088-031f-46ae-977d-c164ed4b237b]
- 21. Advances in Extremophile Research: Biotechnological ... [https://www.mdpi.com/2075-1729/14/9/1205]
- 22. Stability and rigidity/flexibility-two sides of the same coin? [https://pubmed.ncbi.nlm.nih.gov/23416444/]
- 23. Cytoplasmic fluidity and the cold life: proteome stability is ... [https://www.nature.com/articles/s41467-025-65270-5]
- 24. 1GV1: Structural Basis for Thermophilic ... Protein Stability [https://www.rcsb.org/structure/1gv1]
- 25. Next-generation biotechnology inspired by extremes [https://pmc.ncbi.nlm.nih.gov/articles/PMC11893800/]
- 26. Adaption strategies of extremophiles and the construction ... [https://www.sciencedirect.com/science/article/abs/pii/S0013935125012307]
- 27. Genomic and metabolic network properties in thermophiles ... [https://www.nature.com/articles/s41598-025-05030-z]
- 28. Lessons from Extremophiles: Functional Adaptations and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11299111/]
- 29. Understanding activity-stability tradeoffs in biocatalysts by ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10902396/]
- 30. Extremozymes: Challenges and opportunities on the road ... [https://www.sciencedirect.com/science/article/abs/pii/S1359511324001314]
- 31. Precision enzyme discovery through targeted mining of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10781932/]
- 32. Opportunities and challenges of using metagenomic data to ... [https://microbiomejournal.biomedcentral.com/articles/10.1186/s40168-022-01272-5]
- 33. Integrating Metagenomic and Culture-Based Techniques to ... [https://www.mdpi.com/2304-8158/14/3/352]
- 34. Comparison of microbial culture, metagenomic next ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12343261/]
- 35. Benchmarking Metagenomic Pipelines for the Detection of ... [https://www.sciencedirect.com/science/article/pii/S0362028X25001358]
- 36. Metagenomic and Culture-Based Analyses of Microbial ... [https://www.mdpi.com/2079-7737/12/10/1300]
- 37. Approaches to Unmask Functioning of the Uncultured ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2022.845562/full]
- 38. Perspectives on the microorganism of extreme ... [https://www.sciencedirect.com/science/article/pii/S2666517422000311]
- 39. Enzymes from Extreme Environments and their Industrial ... [https://www.frontiersin.org/journals/bioengineering-and-biotechnology/articles/10.3389/fbioe.2015.00161/full]
- 40. Single-Cell Genomics of Microbial Dark Matter [https://pubmed.ncbi.nlm.nih.gov/30298250/]
- 41. Recent Advances in Function-based Metagenomic Screening [https://pmc.ncbi.nlm.nih.gov/articles/PMC6411959/]
- 42. Printing Microbial Dark Matter: Using Single Cell ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2021.635506/full]
- 43. Unraveling the functional dark matter through global ... [https://www.nature.com/articles/s41586-023-06583-7]
- 44. Understanding microbial ecology and evolution with single ... [https://www.nature.com/articles/s41576-025-00918-y]
- 45. Extremophiles: A Novel Source of Industrially Important Enzymes [https://scialert.net/fulltext/?doi=biotech.2011.121.135]
- 46. From the Discovery of Extremozymes to an Enzymatic ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8790482/]
- 47. Strict Regulation of Gene Expression from a High-Copy ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2442401/]
- 48. Extremophiles in a changing world [https://link.springer.com/article/10.1007/s00792-024-01341-7]
- 49. Optimization Strategy of Expression Vectors and ... [https://www.mdpi.com/1422-0067/26/22/10812]
- 50. Editorial: Mining, Designing, Mechanisms and Applications ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2021.709377/full]
- 51. Desirable L-asparaginases for treating cancer and current ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2024.1269282/full]
- 52. Characterization of a Type II L-Asparaginase from the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10672034/]
- 53. A Novel Type II L-Asparaginase from the Halotolerant ... [https://www.preprints.org/manuscript/202309.2166]
- 54. Tapping the untapped: type II L-asparaginase from marine ... [https://www.nature.com/articles/s41598-025-08962-8]
- 55. The potential of halophilic and halotolerant bacteria for the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4908666/]
- 56. Structural insights into the γ -lactamase activity and substrate... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5353710/]
- 57. Scope, limitations and classification of lactamases [https://www.sciencedirect.com/science/article/abs/pii/S0168165616301663]
- 58. Identification and characterization of a novel (+)- γ -lactamase from... [https://link.springer.com/article/10.1007/s00253-016-7643-0]
- 59. Bacterial spore surface display system for enzyme ... [https://www.sciencedirect.com/science/article/abs/pii/S0924224425004121]
- 60. Scaling bioethanol for the future: the commercialization ... [https://www.frontiersin.org/journals/energy-research/articles/10.3389/fenrg.2025.1565273/full]
- 61. Techno-economic assessment of polyhydroxyalkanoates ... [https://www.sciencedirect.com/science/article/abs/pii/S0956053X25002983]
- 62. harnessing the potential of pH-resilient algal strains for ... [https://pubs.rsc.org/en/content/articlehtml/2025/va/d4va00247d]
- 63. Extremophilic Microorganisms as a Source of Emerging ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11716999/]
- 64. recovery from inclusion bodies of Escherichia coli using mild... Protein [https://microbialcellfactories.biomedcentral.com/articles/10.1186/s12934-015-0222-8]
- 65. Codon usage: Nature's roadmap to expression and folding of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3166658/]
- 66. A code within the genetic code: codon usage regulates co ... [https://biosignaling.biomedcentral.com/articles/10.1186/s12964-020-00642-6]
- 67. Exploring Codon Adjustment Strategies towards ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8310279/]
- 68. Chaperone-mediated thermotolerance in ... [https://www.sciencedirect.com/science/article/pii/S2666498425001085?via%3Dihub]
- 69. Beyond directed evolution—semi-rational protein ... [https://www.sciencedirect.com/science/article/abs/pii/S0958166910001540]
- 70. Editorial: Extremozymes: characteristics, structure, protein ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2024.1423463/full]
- 71. Frontiers | Editorial: Industrial Application of Extreme Microbes ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2025.1697504/abstract]
- 72. Enzyme Immobilization Technologies and Industrial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9933091/]
- 73. Review of research progress in immobilization and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12273361/]
- 74. Enzyme immobilization: an overview on techniques and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3563746/]
- 75. Immobilization Techniques - (Physical Chemistry I) [https://fiveable.me/key-terms/physical-chemistry-i/immobilization-techniques]
- 76. Enhancing industrial biocatalyst performance and cost ... [https://www.sciencedirect.com/science/article/abs/pii/S0141813025048305]
- 77. Covalent immobilization: A review from an enzyme ... [https://www.sciencedirect.com/science/article/pii/S1385894724095457]
- 78. Reusability of immobilized β-glucosidase on sodium ... [https://www.sciencedirect.com/science/article/pii/S1319610322000990]
- 79. Reusability of Immobilized Cells for Subsequent Balsamic- ... [https://www.mdpi.com/2311-5637/6/4/103]
- 80. Exploring Principles of Bioreactor Scale-Up [https://www.bioprocessintl.com/bioreactors/lessons-in-bioreactor-scale-up-part-1-mdash-exploring-introductory-principles]
- 81. Tailoring industrial enzymes for thermostability and activity ... [https://www.nature.com/articles/s41467-025-55944-5]
- 82. sciencedirect.com/topics/medicine-and-dentistry/ microbial - enzyme [https://www.sciencedirect.com/topics/medicine-and-dentistry/microbial-enzyme]
- 83. Protein stability engineering insights revealed by domain ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6697890/]
- 84. Accurate predictions of protein mutational effects ... [https://www.nature.com/articles/s42004-025-01771-0]
- 85. Function and biotechnology of extremophilic enzymes in low ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3310334/]
- 86. A live bacteria enzyme assay for identification of human ... [https://www.nature.com/articles/s41551-025-01391-y]
- 87. Exploring enzyme thermostability enhancement through ... [https://www.sciencedirect.com/science/article/abs/pii/S0022286025005381]
- 88. The effects of temperature on the kinetics and stability ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC1219248/]
- 89. Improving the Thermostability and Activity of a ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4542237/]
- 90. Establishment of mesophilic-like catalytic properties in a ... [https://www.nature.com/articles/s41598-019-45560-x]
- 91. Differences in the catalytic mechanisms of mesophilic and ... [https://www.sciencedirect.com/science/article/abs/pii/S0006291X12000435]
- 92. Solvent tolerant enzymes in extremophiles: Adaptations ... [https://www.sciencedirect.com/science/article/abs/pii/S0141813023009455]
- 93. Industrial Biotechnology Based on Enzymes From Extreme ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9036996/]
- 94. A novel salt- and organic solvent-tolerant phosphite ... [https://www.frontiersin.org/journals/bioengineering-and-biotechnology/articles/10.3389/fbioe.2023.1255582/full]
- 95. Extremophile Microbial Communities and Enzymes for Bioenergetic Application Based on Multi-Omics Tools - PMC [https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7521039/]
- 96. Extremophiles Breakthrough: Hot Topics and Current ... [https://www.mdpi.com/2075-1729/15/3/434]
- 97. Frontiers | Cold and Hot Extremozymes: Industrial ... [https://www.frontiersin.org/journals/bioengineering-and-biotechnology/articles/10.3389/fbioe.2015.00148/full]
- 98. Extremozymes and compatible solute production potential ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11231302/]
- 99. Enzyme Engineering: Performance Optimization, Novel ... [https://www.mdpi.com/2304-8158/13/23/3846]
- 100. The Global Sustainable Enzymes Market 2026-2036 [https://www.futuremarketsinc.com/the-global-sustainable-enzymes-market-2026-2036/]